This article contains too many pictures for its overall length. (December 2023) |
Marine life, sea life, or ocean life is the plants, animals, and other organisms that live in the salt water of seas or oceans, or the brackish water of coastal estuaries. At a fundamental level, marine life affects the nature of the planet. Marine organisms, mostly microorganisms, produce oxygen and sequester carbon. Marine life, in part, shape and protect shorelines, and some marine organisms even help create new land (e.g. coral building reefs).


Most life forms evolved initially in marine habitats. By volume, oceans provide about 90% of the living space on the planet.[2] The earliest vertebrates appeared in the form of fish,[3] which live exclusively in water. Some of these evolved into amphibians, which spend portions of their lives in water and portions on land. One group of amphibians evolved into reptiles and mammals and a few subsets of each returned to the ocean as sea snakes, sea turtles, seals, manatees, and whales. Plant forms such as kelp and other algae grow in the water and are the basis for some underwater ecosystems. Plankton forms the general foundation of the ocean food chain, particularly phytoplankton which are key primary producers.
Marine invertebrates exhibit a wide range of modifications to survive in poorly oxygenated waters, including breathing tubes as in mollusc siphons. Fish have gills instead of lungs, although some species of fish, such as the lungfish, have both. Marine mammals (e.g. dolphins, whales, otters, and seals) need to surface periodically to breathe air.
As of 2023[update], more than 242,000 marine species have been documented, and perhaps two million marine species are yet to be documented. An average of 2,332 new species per year are being described.[4][5]
Marine species range in size from the microscopic like phytoplankton, which can be as small as 0.02 micrometres, to huge cetaceans like the blue whale – the largest known animal, reaching 33 m (108 ft) in length.[6][7] Marine microorganisms, including protists and bacteria and their associated viruses, have been variously estimated as constituting about 70%[8] or about 90%[9][1] of the total marine biomass. Marine life is studied scientifically in both marine biology and in biological oceanography. The term marine comes from the Latin mare, meaning "sea" or "ocean".
Water
edit
There is no life without water.[10] It has been described as the universal solvent for its ability to dissolve many substances,[11][12] and as the solvent of life.[13] Water is the only common substance to exist as a solid, liquid, and gas under conditions normal to life on Earth.[14] The Nobel Prize winner Albert Szent-Györgyi referred to water as the mater und matrix: the mother and womb of life.[15]

The abundance of surface water on Earth is a unique feature in the Solar System. Earth's hydrosphere consists chiefly of the oceans but technically includes all water surfaces in the world, including inland seas, lakes, rivers, and underground waters down to a depth of 2,000 metres (6,600 ft). The deepest underwater location is Challenger Deep of the Mariana Trench in the Pacific Ocean, having a depth of 10,900 metres (6.8 mi).[note 1][16]
Conventionally, the planet is divided into five separate oceans, but these oceans all connect into a single world ocean.[17] The mass of this world ocean is 1.35×1018 metric tons or about 1/4400 of Earth's total mass. The world ocean covers an area of 3.618×108 km2 with a mean depth of 3682 m, resulting in an estimated volume of 1.332×109 km3.[18] If all of Earth's crustal surface was at the same elevation as a smooth sphere, the depth of the resulting world ocean would be about 2.7 kilometres (1.7 mi).[19][20]

About 97.5% of the water on Earth is saline; the remaining 2.5% is fresh water. Most fresh water – about 69% – is present as ice in ice caps and glaciers.[21] The average salinity of Earth's oceans is about 35 grams (1.2 oz) of salt per kilogram of seawater (3.5% salt).[22] Most of the salt in the ocean comes from the weathering and erosion of rocks on land.[23] Some salts are released from volcanic activity or extracted from cool igneous rocks.[24]
The oceans are also a reservoir of dissolved atmospheric gases, which are essential for the survival of many aquatic life forms.[25] Sea water has an important influence on the world's climate, with the oceans acting as a large heat reservoir.[26] Shifts in the oceanic temperature distribution can cause significant weather shifts, such as the El Niño-Southern Oscillation.[27]

Altogether the ocean occupies 71 percent of the world surface,[2] averaging nearly 3.7 kilometres (2.3 mi) in depth.[28] By volume, the ocean provides about 90 percent of the living space on the planet.[2] The science fiction writer Arthur C. Clarke has pointed out it would be more appropriate to refer to planet Earth as planet Ocean.[29][30]
However, water is found elsewhere in the Solar System. Europa, one of the moons orbiting Jupiter, is slightly smaller than the Earth's Moon. There is a strong possibility a large saltwater ocean exists beneath its ice surface.[31] It has been estimated the outer crust of solid ice is about 10–30 km (6–19 mi) thick and the liquid ocean underneath is about 100 km (60 mi) deep.[32] This would make Europa's ocean over twice the volume of the Earth's ocean. There has been speculation Europa's ocean could support life,[33][34] and could be capable of supporting multicellular microorganisms if hydrothermal vents are active on the ocean floor.[35] Enceladus, a small icy moon of Saturn, also has what appears to be an underground ocean which actively vents warm water from the moon's surface.[36]
Evolution
editHistorical development
editThe Earth is about 4.54 billion years old.[37][38][39] The earliest undisputed evidence of life on Earth dates from at least 3.5 billion years ago,[40][41] during the Eoarchean era after a geological crust started to solidify following the earlier molten Hadean Eon. Microbial mat fossils have been found in 3.48 billion-year-old sandstone in Western Australia.[42][43] Other early physical evidence of a biogenic substance is graphite in 3.7 billion-year-old metasedimentary rocks discovered in Western Greenland[44] as well as "remains of biotic life" found in 4.1 billion-year-old rocks in Western Australia.[45][46] According to one of the researchers, "If life arose relatively quickly on Earth … then it could be common in the universe."[45]
All organisms on Earth are descended from a common ancestor or ancestral gene pool.[47][48] Highly energetic chemistry is thought to have produced a self-replicating molecule around 4 billion years ago, and half a billion years later the last common ancestor of all life existed.[49] The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions.[50] The beginning of life may have included self-replicating molecules such as RNA[51] and the assembly of simple cells.[52] In 2016 scientists reported a set of 355 genes from the last universal common ancestor (LUCA) of all life, including microorganisms, living on Earth.[53]
Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events.[54] The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits and finally, that organisms can be classified using these similarities into a hierarchy of nested groups—similar to a family tree.[55] However, modern research has suggested that, due to horizontal gene transfer, this "tree of life" may be more complicated than a simple branching tree since some genes have spread independently between distantly related species.[56][57]
Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record.[58] By comparing the anatomies of both modern and extinct species, paleontologists can infer the lineages of those species. However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.

More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and amino acids.[60] The development of molecular genetics has revealed the record of evolution left in organisms' genomes: dating when species diverged through the molecular clock produced by mutations.[61] For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 98% of their genomes and analysing the few areas where they differ helps shed light on when the common ancestor of these species existed.[62]
Prokaryotes inhabited the Earth from approximately 3–4 billion years ago.[63][64] No obvious changes in morphology or cellular organisation occurred in these organisms over the next few billion years.[65] The eukaryotic cells emerged between 1.6 and 2.7 billion years ago. The next major change in cell structure came when bacteria were engulfed by eukaryotic cells, in a cooperative association called endosymbiosis.[66][67] The engulfed bacteria and the host cell then underwent coevolution, with the bacteria evolving into either mitochondria or hydrogenosomes.[68] Another engulfment of cyanobacterial-like organisms led to the formation of chloroplasts in algae and plants.[69]

The history of life was that of the unicellular eukaryotes, prokaryotes and archaea until about 610 million years ago when multicellular organisms began to appear in the oceans in the Ediacaran period.[63][70] The evolution of multicellularity occurred in multiple independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime moulds and myxobacteria.[71] In 2016 scientists reported that, about 800 million years ago, a minor genetic change in a single molecule called GK-PID may have allowed organisms to go from a single cell organism to one of many cells.[72]
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over a span of about 10 million years, in an event called the Cambrian explosion. Here, the majority of types of modern animals appeared in the fossil record, as well as unique lineages that subsequently became extinct.[73] Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere from photosynthesis.[74]
About 500 million years ago, plants and fungi started colonising the land. Evidence for the appearance of the first land plants occurs in the Ordovician, around 450 million years ago, in the form of fossil spores.[75] Land plants began to diversify in the Late Silurian, from around 430 million years ago.[76] The colonisation of the land by plants was soon followed by arthropods and other animals.[77] Insects were particularly successful and even today make up the majority of animal species.[78] Amphibians first appeared around 364 million years ago, followed by early amniotes and birds around 155 million years ago (both from "reptile"-like lineages), mammals around 129 million years ago, homininae around 10 million years ago and modern humans around 250,000 years ago.[79][80][81] However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to be highly successful and dominate the Earth, with the majority of both biomass and species being prokaryotes.[82]
Estimates on the number of Earth's current species range from 10 million to 14 million,[83] of which about 1.2 million have been documented and over 86 percent have not yet been described.[84]
Microorganisms
edit

Microorganisms make up about 70% of the marine biomass.[8] A microorganism, or microbe, is a microscopic organism too small to be recognised with the naked eye. It can be single-celled[85] or multicellular. Microorganisms are diverse and include all bacteria and archaea, most protozoa such as algae, fungi, and certain microscopic animals such as rotifers.
Many macroscopic animals and plants have microscopic juvenile stages. Some microbiologists also classify viruses (and viroids) as microorganisms, but others consider these as nonliving.[86][87]
Microorganisms are crucial to nutrient recycling in ecosystems as they act as decomposers. Some microorganisms are pathogenic, causing disease and even death in plants and animals.[88] As inhabitants of the largest environment on Earth, microbial marine systems drive changes in every global system. Microbes are responsible for virtually all the photosynthesis that occurs in the ocean, as well as the cycling of carbon, nitrogen, phosphorus, other nutrients and trace elements.[89]

_-_Stephanopyxis_sp._-_630x_(14117161428).jpg)

Microscopic life undersea is diverse and still poorly understood, such as for the role of viruses in marine ecosystems.[90] Most marine viruses are bacteriophages, which are harmless to plants and animals, but are essential to the regulation of saltwater and freshwater ecosystems.[91]: 5 They infect and destroy bacteria in aquatic microbial communities, and are the most important mechanism of recycling carbon in the marine environment. The organic molecules released from the dead bacterial cells stimulate fresh bacterial and algal growth.[91]: 593 Viral activity may also contribute to the biological pump, the process whereby carbon is sequestered in the deep ocean.[92]


A stream of airborne microorganisms circles the planet above weather systems but below commercial air lanes.[93] Some peripatetic microorganisms are swept up from terrestrial dust storms, but most originate from marine microorganisms in sea spray. In 2018, scientists reported that hundreds of millions of viruses and tens of millions of bacteria are deposited daily on every square meter around the planet.[94][95]
Microscopic organisms live throughout the biosphere. The mass of prokaryote microorganisms — which includes bacteria and archaea, but not the nucleated eukaryote microorganisms — may be as much as 0.8 trillion tons of carbon (of the total biosphere mass, estimated at between 1 and 4 trillion tons).[96] Single-celled barophilic marine microbes have been found at a depth of 10,900 m (35,800 ft) in the Mariana Trench, the deepest spot in the Earth's oceans.[97][98] Microorganisms live inside rocks 580 m (1,900 ft) below the sea floor under 2,590 m (8,500 ft) of ocean off the coast of the northwestern United States,[97][99] as well as 2,400 m (7,900 ft; 1.5 mi) beneath the seabed off Japan.[100] The greatest known temperature at which microbial life can exist is 122 °C (252 °F) (Methanopyrus kandleri).[101] In 2014, scientists confirmed the existence of microorganisms living 800 m (2,600 ft) below the ice of Antarctica.[102][103] According to one researcher, "You can find microbes everywhere — they're extremely adaptable to conditions, and survive wherever they are."[97]
Marine viruses
editViruses are small infectious agents that do not have their own metabolism and can replicate only inside the living cells of other organisms.[104] Viruses can infect all types of life forms, from animals and plants to microorganisms, including bacteria and archaea.[105] The linear size of the average virus is about one one-hundredth that of the average bacterium. Most viruses cannot be seen with an optical microscope so electron microscopes are used instead.[106]
Viruses are found wherever there is life and have probably existed since living cells first evolved.[107] The origin of viruses is unclear because they do not form fossils, so molecular techniques have been used to compare the DNA or RNA of viruses and are a useful means of investigating how they arise.[108]
Viruses are now recognised as ancient and as having origins that pre-date the divergence of life into the three domains.[109] But the origins of viruses in the evolutionary history of life are unclear: some may have evolved from plasmids—pieces of DNA that can move between cells—while others may have evolved from bacteria. In evolution, viruses are an important means of horizontal gene transfer, which increases genetic diversity.[110]



Opinions differ on whether viruses are a form of life or organic structures that interact with living organisms.[111] They are considered by some to be a life form, because they carry genetic material, reproduce by creating multiple copies of themselves through self-assembly, and evolve through natural selection. However they lack key characteristics such as a cellular structure generally considered necessary to count as life. Because they possess some but not all such qualities, viruses have been described as replicators[111] and as "organisms at the edge of life".[112]

Bacteriophages, often just called phages, are viruses that parasite bacteria and archaea. Marine phages parasite marine bacteria and archaea, such as cyanobacteria.[113] They are a common and diverse group of viruses and are the most abundant biological entity in marine environments, because their hosts, bacteria, are typically the numerically dominant cellular life in the sea. Generally there are about 1 million to 10 million viruses in each mL of seawater, or about ten times more double-stranded DNA viruses than there are cellular organisms,[114][115] although estimates of viral abundance in seawater can vary over a wide range.[116][117] Tailed bacteriophages appear to dominate marine ecosystems in number and diversity of organisms.[113] Bacteriophages belonging to the families Corticoviridae,[118] Inoviridae[119] and Microviridae[120] are also known to infect diverse marine bacteria.
Microorganisms make up about 70% of the marine biomass.[8] It is estimated viruses kill 20% of this biomass each day and that there are 15 times as many viruses in the oceans as there are bacteria and archaea. Viruses are the main agents responsible for the rapid destruction of harmful algal blooms,[115] which often kill other marine life.[121] The number of viruses in the oceans decreases further offshore and deeper into the water, where there are fewer host organisms.[92]
There are also archaeal viruses which replicate within archaea: these are double-stranded DNA viruses with unusual and sometimes unique shapes.[122][123] These viruses have been studied in most detail in the thermophilic archaea, particularly the orders Sulfolobales and Thermoproteales.[124]
Viruses are an important natural means of transferring genes between different species, which increases genetic diversity and drives evolution.[110] It is thought that viruses played a central role in the early evolution, before the diversification of bacteria, archaea and eukaryotes, at the time of the last universal common ancestor of life on Earth.[125] Viruses are still one of the largest reservoirs of unexplored genetic diversity on Earth.[92]
Marine bacteria
edit

Bacteria constitute a large domain of prokaryotic microorganisms. Typically a few micrometres in length, bacteria have a number of shapes, ranging from spheres to rods and spirals. Bacteria were among the first life forms to appear on Earth, and are present in most of its habitats. Bacteria inhabit soil, water, acidic hot springs, radioactive waste,[126] and the deep portions of Earth's crust. Bacteria also live in symbiotic and parasitic relationships with plants and animals.
Once regarded as plants constituting the class Schizomycetes, bacteria are now classified as prokaryotes. Unlike cells of animals and other eukaryotes, bacterial cells do not contain a nucleus and rarely harbour membrane-bound organelles. Although the term bacteria traditionally included all prokaryotes, the scientific classification changed after the discovery in the 1990s that prokaryotes consist of two very different groups of organisms that evolved from an ancient common ancestor. These evolutionary domains are called Bacteria and Archaea.[127]
The ancestors of modern bacteria were unicellular microorganisms that were the first forms of life to appear on Earth, about 4 billion years ago. For about 3 billion years, most organisms were microscopic, and bacteria and archaea were the dominant forms of life.[65][128] Although bacterial fossils exist, such as stromatolites, their lack of distinctive morphology prevents them from being used to examine the history of bacterial evolution, or to date the time of origin of a particular bacterial species. However, gene sequences can be used to reconstruct the bacterial phylogeny, and these studies indicate that bacteria diverged first from the archaeal/eukaryotic lineage.[129] Bacteria were also involved in the second great evolutionary divergence, that of the archaea and eukaryotes. Here, eukaryotes resulted from the entering of ancient bacteria into endosymbiotic associations with the ancestors of eukaryotic cells, which were themselves possibly related to the Archaea.[67][66] This involved the engulfment by proto-eukaryotic cells of alphaproteobacterial symbionts to form either mitochondria or hydrogenosomes, which are still found in all known Eukarya. Later on, some eukaryotes that already contained mitochondria also engulfed cyanobacterial-like organisms. This led to the formation of chloroplasts in algae and plants. There are also some algae that originated from even later endosymbiotic events. Here, eukaryotes engulfed a eukaryotic algae that developed into a "second-generation" plastid.[130][131] This is known as secondary endosymbiosis.
-
 The marine Thiomargarita namibiensis, the largest known bacterium
The marine Thiomargarita namibiensis, the largest known bacterium -

-
![The chloroplasts of glaucophytes have a peptidoglycan layer, evidence suggesting their endosymbiotic origin from cyanobacteria.[132]](//upload.wikimedia.org/wikipedia/commons/thumb/c/c9/Glaucocystis_sp.jpg/280px-Glaucocystis_sp.jpg) The chloroplasts of glaucophytes have a peptidoglycan layer, evidence suggesting their endosymbiotic origin from cyanobacteria.[132]
The chloroplasts of glaucophytes have a peptidoglycan layer, evidence suggesting their endosymbiotic origin from cyanobacteria.[132] -
 Bacteria can be beneficial. This Pompeii worm, an extremophile found only at hydrothermal vents, has a protective cover of bacteria.
Bacteria can be beneficial. This Pompeii worm, an extremophile found only at hydrothermal vents, has a protective cover of bacteria.


![The chloroplasts of glaucophytes have a peptidoglycan layer, evidence suggesting their endosymbiotic origin from cyanobacteria.[132]](/wiki/File:Glaucocystis_sp.jpg)

The largest known bacterium, the marine Thiomargarita namibiensis, can be visible to the naked eye and sometimes attains 0.75 mm (750 μm).[133][134]
Marine archaea
edit
The archaea (Greek for ancient[136]) constitute a domain and kingdom of single-celled microorganisms. These microbes are prokaryotes, meaning they have no cell nucleus or any other membrane-bound organelles in their cells.
Archaea were initially classified as bacteria, but this classification is outdated.[137] Archaeal cells have unique properties separating them from the other two domains of life, Bacteria and Eukaryota. The Archaea are further divided into multiple recognized phyla. Classification is difficult because the majority have not been isolated in the laboratory and have only been detected by analysis of their nucleic acids in samples from their environment.
Archaea and bacteria are generally similar in size and shape, although a few archaea have very strange shapes, such as the flat and square-shaped cells of Haloquadratum walsbyi.[138] Despite this morphological similarity to bacteria, archaea possess genes and several metabolic pathways that are more closely related to those of eukaryotes, notably the enzymes involved in transcription and translation. Other aspects of archaeal biochemistry are unique, such as their reliance on ether lipids in their cell membranes, such as archaeols. Archaea use more energy sources than eukaryotes: these range from organic compounds, such as sugars, to ammonia, metal ions or even hydrogen gas. Salt-tolerant archaea (the Haloarchaea) use sunlight as an energy source, and other species of archaea fix carbon; however, unlike plants and cyanobacteria, no known species of archaea does both. Archaea reproduce asexually by binary fission, fragmentation, or budding; unlike bacteria and eukaryotes, no known species forms spores.
Archaea are particularly numerous in the oceans, and the archaea in plankton may be one of the most abundant groups of organisms on the planet. Archaea are a major part of Earth's life and may play roles in both the carbon cycle and the nitrogen cycle.
-
 Halobacteria, found in water near saturated with salt, are now recognised as archaea.
Halobacteria, found in water near saturated with salt, are now recognised as archaea. -
 Flat, square-shaped cells of the archaea Haloquadratum walsbyi
Flat, square-shaped cells of the archaea Haloquadratum walsbyi -
 Methanosarcina barkeri, a marine archaea that produces methane
Methanosarcina barkeri, a marine archaea that produces methane -
 Thermophiles, such as Pyrolobus fumarii, survive well over 100 °C.
Thermophiles, such as Pyrolobus fumarii, survive well over 100 °C. -
 Drawing of another marine thermophile, Pyrococcus furiosus
Drawing of another marine thermophile, Pyrococcus furiosus





Marine protists
editProtists are eukaryotes that cannot be classified as plants, fungi or animals. They are usually single-celled and microscopic. Life originated as single-celled prokaryotes (bacteria and archaea) and later evolved into more complex eukaryotes. Eukaryotes are the more developed life forms known as plants, animals, fungi and protists. The term protist came into use historically as a term of convenience for eukaryotes that cannot be strictly classified as plants, animals or fungi. They are not a part of modern cladistics, because they are paraphyletic (lacking a common ancestor). Protists can be broadly divided into four groups depending on whether their nutrition is plant-like, animal-like, fungus-like,[139] or a mixture of these.[140]
Protists according to how they get food
| |||||||
|---|---|---|---|---|---|---|---|
| Type of protist | Description | Example | Other examples | ||||
| Plant-like | Autotrophic protists that make their own food without needing to consume other organisms, usually by using photosynthesis | 
|
Red algae, Cyanidium sp. | Green algae, brown algae, diatoms and some dinoflagellates. Plant-like protists are important components of phytoplankton discussed below. | |||
| Animal-like | Heterotrophic protists that get their food consuming other organisms | 
|
Radiolarian protist as drawn by Haeckel | Foraminiferans, and some marine amoebae, ciliates and flagellates. | |||
| Fungus-like | Saprotrophic protists that get their food from the remains of organisms that have broken down and decayed | 
|
Marine slime nets form labyrinthine networks of tubes in which amoeba without pseudopods can travel | Marine lichen | |||
| Mixotropes | Various
|
Mixotrophic and osmotrophic protists that get their food from a combination of the above | _(cropped).jpg)
|
Euglena mutabilis, a photosynthetic flagellate | Many marine mixotropes are found among protists, including among ciliates, Rhizaria and dinoflagellates [141] | ||


Protists are highly diverse organisms currently organised into 18 phyla, but are not easy to classify.[143][144] Studies have shown high protist diversity exists in oceans, deep sea-vents and river sediments, suggesting a large number of eukaryotic microbial communities have yet to be discovered.[145][146] There has been little research on mixotrophic protists, but recent studies in marine environments found mixotrophic protests contribute a significant part of the protist biomass.[141]
- Single-celled and microscopic protists
-
![Diatoms are a major algae group generating about 20% of world oxygen production.[147]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
-
![Diatoms have glass like cell walls made of silica and called frustules.[148]](//upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)
-
 Fossil diatom frustule from 32 to 40 mya
Fossil diatom frustule from 32 to 40 mya -

-
 Single-celled alga, Gephyrocapsa oceanica
Single-celled alga, Gephyrocapsa oceanica -
 Two dinoflagellates
Two dinoflagellates -
 Zooxanthellae is a photosynthetic algae that lives inside hosts like coral.
Zooxanthellae is a photosynthetic algae that lives inside hosts like coral. -

-

-
 This ciliate is digesting cyanobacteria. The cytostome or mouth is at the bottom right.
This ciliate is digesting cyanobacteria. The cytostome or mouth is at the bottom right.
![Diatoms are a major algae group generating about 20% of world oxygen production.[147]](/wiki/File:Diatoms_through_the_microscope.jpg)
![Diatoms have glass like cell walls made of silica and called frustules.[148]](/wiki/File:Diatom_algae_Amphora_sp.jpg)

.jpg)
.jpg)





In contrast to the cells of prokaryotes, the cells of eukaryotes are highly organised. Plants, animals and fungi are usually multi-celled and are typically macroscopic. Most protists are single-celled and microscopic. But there are exceptions. Some single-celled marine protists are macroscopic. Some marine slime molds have unique life cycles that involve switching between unicellular, colonial, and multicellular forms.[149] Other marine protist are neither single-celled nor microscopic, such as seaweed.
- Macroscopic protists (see also unicellular macroalgae →)
-
 The single-celled giant amoeba has up to 1000 nuclei and reaches lengths of 5 mm (0.20 in).
The single-celled giant amoeba has up to 1000 nuclei and reaches lengths of 5 mm (0.20 in). -
![Gromia sphaerica is a large spherical testate amoeba which makes mud trails. Its diameter is up to 3.8 cm (1.5 in).[150]](//upload.wikimedia.org/wikipedia/commons/thumb/9/99/Gromia_in_situ_closeup.png/266px-Gromia_in_situ_closeup.png) Gromia sphaerica is a large spherical testate amoeba which makes mud trails. Its diameter is up to 3.8 cm (1.5 in).[150]
Gromia sphaerica is a large spherical testate amoeba which makes mud trails. Its diameter is up to 3.8 cm (1.5 in).[150] -
 Spiculosiphon oceana, a unicellular foraminiferan with an appearance and lifestyle that mimics a sponge, grows to 5 cm long.
Spiculosiphon oceana, a unicellular foraminiferan with an appearance and lifestyle that mimics a sponge, grows to 5 cm long. -
![The xenophyophore, another single-celled foraminiferan, lives in abyssal zones. It has a giant shell up to 20 cm (7.9 in) across.[151]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5d/Xenophyophore.jpg/292px-Xenophyophore.jpg) The xenophyophore, another single-celled foraminiferan, lives in abyssal zones. It has a giant shell up to 20 cm (7.9 in) across.[151]
The xenophyophore, another single-celled foraminiferan, lives in abyssal zones. It has a giant shell up to 20 cm (7.9 in) across.[151] -
 Giant kelp, a brown algae, is not a true plant, yet it is multicellular and can grow to 50m.
Giant kelp, a brown algae, is not a true plant, yet it is multicellular and can grow to 50m.

![Gromia sphaerica is a large spherical testate amoeba which makes mud trails. Its diameter is up to 3.8 cm (1.5 in).[150]](/wiki/File:Gromia_in_situ_closeup.png)

![The xenophyophore, another single-celled foraminiferan, lives in abyssal zones. It has a giant shell up to 20 cm (7.9 in) across.[151]](/wiki/File:Xenophyophore.jpg)

Protists have been described as a taxonomic grab bag where anything that doesn't fit into one of the main biological kingdoms can be placed.[152] Some modern authors prefer to exclude multicellular organisms from the traditional definition of a protist, restricting protists to unicellular organisms.[153][154] This more constrained definition excludes seaweeds and slime molds.[155]
Marine microanimals
edit| External videos | |
|---|---|
As juveniles, animals develop from microscopic stages, which can include spores, eggs and larvae. At least one microscopic animal group, the parasitic cnidarian Myxozoa, is unicellular in its adult form, and includes marine species. Other adult marine microanimals are multicellular. Microscopic adult arthropods are more commonly found inland in freshwater, but there are marine species as well. Microscopic adult marine crustaceans include some copepods, cladocera and tardigrades (water bears). Some marine nematodes and rotifers are also too small to be recognised with the naked eye, as are many loricifera, including the recently discovered anaerobic species that spend their lives in an anoxic environment.[156][157] Copepods contribute more to the secondary productivity and carbon sink of the world oceans than any other group of organisms.[158][159] While mites are not normally thought of as marine organisms, most species of the family Halacaridae live in the sea.[160]
- Marine microanimals
-
 Over 10,000 marine species are copepods, small, often microscopic crustaceans
Over 10,000 marine species are copepods, small, often microscopic crustaceans -
 Darkfield photo of a gastrotrich, a worm-like animal living between sediment particles
Darkfield photo of a gastrotrich, a worm-like animal living between sediment particles -
 Armoured Pliciloricus enigmaticus, about 0.2 mm long, live in spaces between marine gravel.
Armoured Pliciloricus enigmaticus, about 0.2 mm long, live in spaces between marine gravel. -
 Drawing of a tardigrade (water bear) on a grain of sand
Drawing of a tardigrade (water bear) on a grain of sand -
 Rotifers, usually 0.1–0.5 mm long, may look like protists but have many cells and belongs to the Animalia.
Rotifers, usually 0.1–0.5 mm long, may look like protists but have many cells and belongs to the Animalia.




_-_160x_(13402418244).jpg)
Fungi
edit

Over 1500 species of fungi are known from marine environments.[161] These are parasitic on marine algae or animals, or are saprobes feeding on dead organic matter from algae, corals, protozoan cysts, sea grasses, wood and other substrata.[162] Spores of many species have special appendages which facilitate attachment to the substratum.[163] Marine fungi can also be found in sea foam and around hydrothermal areas of the ocean.[164] A diverse range of unusual secondary metabolites is produced by marine fungi.[165]
Mycoplankton are saprotropic members of the plankton communities of marine and freshwater ecosystems.[166][167] They are composed of filamentous free-living fungi and yeasts associated with planktonic particles or phytoplankton.[168] Similar to bacterioplankton, these aquatic fungi play a significant role in heterotrophic mineralization and nutrient cycling.[169] Mycoplankton can be up to 20 mm in diameter and over 50 mm in length.[170]
A typical milliliter of seawater contains about 103 to 104 fungal cells.[171] This number is greater in coastal ecosystems and estuaries due to nutritional runoff from terrestrial communities. A higher diversity of mycoplankton is found around coasts and in surface waters down to 1000 metres, with a vertical profile that depends on how abundant phytoplankton is.[172][173] This profile changes between seasons due to changes in nutrient availability.[174] Marine fungi survive in a constant oxygen deficient environment, and therefore depend on oxygen diffusion by turbulence and oxygen generated by photosynthetic organisms.[175]
Marine fungi can be classified as:[175]
- Lower fungi - adapted to marine habitats (zoosporic fungi, including mastigomycetes: oomycetes and chytridiomycetes)
- Higher fungi - filamentous, modified to planktonic lifestyle (hyphomycetes, ascomycetes, basidiomycetes). Most mycoplankton species are higher fungi.[172]
Lichens are mutualistic associations between a fungus, usually an ascomycete, and an alga or a cyanobacterium. Several lichens are found in marine environments.[176] Many more occur in the splash zone, where they occupy different vertical zones depending on how tolerant they are to submersion.[177] Some lichens live a long time; one species has been dated at 8,600 years.[178] However their lifespan is difficult to measure because what defines the same lichen is not precise.[179] Lichens grow by vegetatively breaking off a piece, which may or may not be defined as the same lichen, and two lichens of different ages can merge, raising the issue of whether it is the same lichen.[179] The sea snail Littoraria irrorata damages plants of Spartina in the sea marshes where it lives, which enables spores of intertidal ascomycetous fungi to colonise the plant. The snail then eats the fungal growth in preference to the grass itself.[180]
According to fossil records, fungi date back to the late Proterozoic era 900-570 million years ago. Fossil marine lichens 600 million years old have been discovered in China.[181] It has been hypothesized that mycoplankton evolved from terrestrial fungi, likely in the Paleozoic era (390 million years ago).[182]
Origin of animals
edit
The earliest animals were marine invertebrates, that is, vertebrates came later. Animals are multicellular eukaryotes,[note 2] and are distinguished from plants, algae, and fungi by lacking cell walls.[183] Marine invertebrates are animals that inhabit a marine environment apart from the vertebrate members of the chordate phylum; invertebrates lack a vertebral column. Some have evolved a shell or a hard exoskeleton.
The earliest animal fossils may belong to the genus Dickinsonia,[184] 571 million to 541 million years ago.[185] Individual Dickinsonia typically resemble a bilaterally symmetrical ribbed oval. They kept growing until they were covered with sediment or otherwise killed,[186] and spent most of their lives with their bodies firmly anchored to the sediment.[187] Their taxonomic affinities are presently unknown, but their mode of growth is consistent with a bilaterian affinity.[188]
Apart from Dickinsonia, the earliest widely accepted animal fossils are the rather modern-looking cnidarians (the group that includes coral, jellyfish, sea anemones and Hydra), possibly from around 580 Ma[189] The Ediacara biota, which flourished for the last 40 million years before the start of the Cambrian,[190] were the first animals more than a very few centimetres long. Like Dickinsonia, many were flat with a "quilted" appearance, and seemed so strange that there was a proposal to classify them as a separate kingdom, Vendozoa.[191] Others, however, have been interpreted as early molluscs (Kimberella[192][193]), echinoderms (Arkarua[194]), and arthropods (Spriggina,[195] Parvancorina[196]). There is still debate about the classification of these specimens, mainly because the diagnostic features which allow taxonomists to classify more recent organisms, such as similarities to living organisms, are generally absent in the Ediacarans. However, there seems little doubt that Kimberella was at least a triploblastic bilaterian animal, in other words, an animal significantly more complex than the cnidarians.[197]
Small shelly fauna are a very mixed collection of fossils found between the Late Ediacaran and Middle Cambrian periods. The earliest, Cloudina, shows signs of successful defense against predation and may indicate the start of an evolutionary arms race. Some tiny Early Cambrian shells almost certainly belonged to molluscs, while the owners of some "armor plates," Halkieria and Microdictyon, were eventually identified when more complete specimens were found in Cambrian lagerstätten that preserved soft-bodied animals.[198]
Body plans and phyla
edit
Invertebrates are grouped into different phyla. Informally phyla can be thought of as a way of grouping organisms according to their body plan.[199][200]: 33 A body plan refers to a blueprint which describes the shape or morphology of an organism, such as its symmetry, segmentation and the disposition of its appendages. The idea of body plans originated with vertebrates, which were grouped into one phylum. But the vertebrate body plan is only one of many, and invertebrates consist of many phyla or body plans. The history of the discovery of body plans can be seen as a movement from a worldview centred on vertebrates, to seeing the vertebrates as one body plan among many. Among the pioneering zoologists, Linnaeus identified two body plans outside the vertebrates; Cuvier identified three; and Haeckel had four, as well as the Protista with eight more, for a total of twelve. For comparison, the number of phyla recognised by modern zoologists has risen to 35.[200]


Historically body plans were thought of as having evolved rapidly during the Cambrian explosion,[204] but a more nuanced understanding of animal evolution suggests a gradual development of body plans throughout the early Palaeozoic and beyond.[205] More generally a phylum can be defined in two ways: as described above, as a group of organisms with a certain degree of morphological or developmental similarity (the phenetic definition), or a group of organisms with a certain degree of evolutionary relatedness (the phylogenetic definition).[205]
In the 1970s there was already a debate about whether the emergence of the modern phyla was "explosive" or gradual but hidden by the shortage of Precambrian animal fossils.[198] A re-analysis of fossils from the Burgess Shale lagerstätte increased interest in the issue when it revealed animals, such as Opabinia, which did not fit into any known phylum. At the time these were interpreted as evidence that the modern phyla had evolved very rapidly in the Cambrian explosion and that the Burgess Shale's "weird wonders" showed that the Early Cambrian was a uniquely experimental period of animal evolution.[206] Later discoveries of similar animals and the development of new theoretical approaches led to the conclusion that many of the "weird wonders" were evolutionary "aunts" or "cousins" of modern groups[207]—for example that Opabinia was a member of the lobopods, a group which includes the ancestors of the arthropods, and that it may have been closely related to the modern tardigrades.[208] Nevertheless, there is still much debate about whether the Cambrian explosion was really explosive and, if so, how and why it happened and why it appears unique in the history of animals.[209]
Earliest animals
editThe deepest-branching animals — the earliest animals that appeared during evolution — are marine non-vertebrate organisms. The earliest animal phyla are the Porifera, Ctenophora, Placozoa and Cnidaria. No member of these clades exhibit body plans with bilateral symmetry.
| ||||||||||||||||||||||||||||||||||
.jpg)

Marine sponges
edit.jpg)
Sponges are animals of the phylum Porifera (from Modern Latin for bearing pores[213]). They are multicellular organisms that have bodies full of pores and channels allowing water to circulate through them, consisting of jelly-like mesohyl sandwiched between two thin layers of cells. They have unspecialized cells that can transform into other types and that often migrate between the main cell layers and the mesohyl in the process. Sponges do not have nervous, digestive or circulatory systems. Instead, most rely on maintaining a constant water flow through their bodies to obtain food and oxygen and to remove wastes.
Sponges are similar to other animals in that they are multicellular, heterotrophic, lack cell walls and produce sperm cells. Unlike other animals, they lack true tissues and organs, and have no body symmetry. The shapes of their bodies are adapted for maximal efficiency of water flow through the central cavity, where it deposits nutrients, and leaves through a hole called the osculum. Many sponges have internal skeletons of spongin and/or spicules of calcium carbonate or silicon dioxide. All sponges are sessile aquatic animals. Although there are freshwater species, the great majority are marine (salt water) species, ranging from tidal zones to depths exceeding 8,800 m (5.5 mi). Some sponges live to great ages; there is evidence of the deep-sea glass sponge Monorhaphis chuni living about 11,000 years.[214][215]
While most of the approximately 5,000–10,000 known species feed on bacteria and other food particles in the water, some host photosynthesizing micro-organisms as endosymbionts and these alliances often produce more food and oxygen than they consume. A few species of sponge that live in food-poor environments have become carnivores that prey mainly on small crustaceans.[216]
-
 Sponge biodiversity. There are four sponge species in this photo.
Sponge biodiversity. There are four sponge species in this photo. -

-
 Venus' flower basket at a depth of 2572 meters
Venus' flower basket at a depth of 2572 meters -

-
 The long-living Monorhaphis chuni
The long-living Monorhaphis chuni

.jpg)
.jpg)
.jpg)

Linnaeus mistakenly identified sponges as plants in the order Algae.[217] For a long time thereafter sponges were assigned to a separate subkingdom, Parazoa (meaning beside the animals).[218] They are now classified as a paraphyletic phylum from which the higher animals have evolved.[219]
Ctenophores
editCtenophores (from Greek for carrying a comb), commonly known as comb jellies, are a phylum that live worldwide in marine waters. They are the largest non-colonial animals to swim with the help of cilia (hairs or combs).[220] Coastal species need to be tough enough to withstand waves and swirling sediment, but some oceanic species are so fragile and transparent that it is very difficult to capture them intact for study.[221] In the past ctenophores were thought to have only a modest presence in the ocean, but it is now known they are often significant and even dominant parts of the planktonic biomass.[222]: 269
The phylum has about 150 known species with a wide range of body forms. Sizes range from a few millimeters to 1.5 m (4 ft 11 in). Cydippids are egg-shaped with their cilia arranged in eight radial comb rows, and deploy retractable tentacles for capturing prey. The benthic platyctenids are generally combless and flat. The coastal beroids have gaping mouths and lack tentacles. Most adult ctenophores prey on microscopic larvae and rotifers and small crustaceans but beroids prey on other ctenophores.
-
 Light diffracting along the comb rows of a cydippid, left tentacle deployed, right retracted
Light diffracting along the comb rows of a cydippid, left tentacle deployed, right retracted -
 Deep-sea ctenophore trailing tentacles studded with tentilla (sub-tentacles)
Deep-sea ctenophore trailing tentacles studded with tentilla (sub-tentacles) -
 Egg-shaped cydippid ctenophore
Egg-shaped cydippid ctenophore -
 Group of small benthic creeping comb jellies streaming tentacles and living symbiotically on a starfish.
Group of small benthic creeping comb jellies streaming tentacles and living symbiotically on a starfish. -
 Lobata sp. with paired thick lobes
Lobata sp. with paired thick lobes -
![The sea walnut has a transient anus which forms only when it needs to defecate.[223]](//upload.wikimedia.org/wikipedia/commons/thumb/f/f8/Mnemiopsis_leidyi_2.jpg/255px-Mnemiopsis_leidyi_2.jpg) The sea walnut has a transient anus which forms only when it needs to defecate.[223]
The sea walnut has a transient anus which forms only when it needs to defecate.[223]



_on_Echniaster_luzonicus_(Seastar).jpg)

![The sea walnut has a transient anus which forms only when it needs to defecate.[223]](/wiki/File:Mnemiopsis_leidyi_2.jpg)

Early writers combined ctenophores with cnidarians. Ctenophores resemble cnidarians in relying on water flow through the body cavity for both digestion and respiration, as well as in having a decentralized nerve net rather than a brain. Also like cnidarians, the bodies of ctenophores consist of a mass of jelly, with one layer of cells on the outside and another lining the internal cavity. In ctenophores, however, these layers are two cells deep, while those in cnidarians are only a single cell deep. While cnidarians exhibit radial symmetry, ctenophores have two anal canals which exhibit biradial symmetry (half-turn rotational symmetry).[224] The position of the ctenophores in the evolutionary family tree of animals has long been debated, and the majority view at present, based on molecular phylogenetics, is that cnidarians and bilaterians are more closely related to each other than either is to ctenophores.[222]: 222
| External videos | |
|---|---|
Placozoa
editPlacozoa (from Greek for flat animals) have the simplest structure of all animals. They are a basal form of free-living (non-parasitic) multicellular organism[225] that do not yet have a common name.[226] They live in marine environments and form a phylum containing so far only three described species, of which the first, the classical Trichoplax adhaerens, was discovered in 1883.[227] Two more species have been discovered since 2017,[228][229] and genetic methods indicate this phylum has a further 100 to 200 undescribed species.[230]



Trichoplax is a small, flattened, animal about one mm across and usually about 25 μm thick. Like the amoebae they superficially resemble, they continually change their external shape. In addition, spherical phases occasionally form which may facilitate movement. Trichoplax lacks tissues and organs. There is no manifest body symmetry, so it is not possible to distinguish anterior from posterior or left from right. It is made up of a few thousand cells of six types in three distinct layers.[231] The outer layer of simple epithelial cells bear cilia which the animal uses to help it creep along the seafloor.[232] Trichoplax feed by engulfing and absorbing food particles – mainly microbes and organic detritus – with their underside.
Marine cnidarians
edit_999_(30695685804).jpg)
Cnidarians (from Greek for nettle) are distinguished by the presence of stinging cells, specialized cells that they use mainly for capturing prey. Cnidarians include corals, sea anemones, jellyfish and hydrozoans. They form a phylum containing over 10,000[233] species of animals found exclusively in aquatic (mainly marine) environments. Their bodies consist of mesoglea, a non-living jelly-like substance, sandwiched between two layers of epithelium that are mostly one cell thick. They have two basic body forms: swimming medusae and sessile polyps, both of which are radially symmetrical with mouths surrounded by tentacles that bear cnidocytes. Both forms have a single orifice and body cavity that are used for digestion and respiration.
Fossil cnidarians have been found in rocks formed about 580 million years ago. Fossils of cnidarians that do not build mineralized structures are rare. Scientists currently think cnidarians, ctenophores and bilaterians are more closely related to calcareous sponges than these are to other sponges, and that anthozoans are the evolutionary "aunts" or "sisters" of other cnidarians, and the most closely related to bilaterians.
Cnidarians are the simplest animals in which the cells are organised into tissues.[234] The starlet sea anemone is used as a model organism in research.[235] It is easy to care for in the laboratory and a protocol has been developed which can yield large numbers of embryos on a daily basis.[236] There is a remarkable degree of similarity in the gene sequence conservation and complexity between the sea anemone and vertebrates.[236] In particular, genes concerned in the formation of the head in vertebrates are also present in the anemone.[237][238]
-
 Sea anemones are common in tidepools.
Sea anemones are common in tidepools. -
 Their tentacles sting and paralyse small fish.
Their tentacles sting and paralyse small fish. -

-
 If an island sinks below the sea, coral growth can keep up with rising water and form an atoll.
If an island sinks below the sea, coral growth can keep up with rising water and form an atoll. -
![The mantle of the red paper lantern jellyfish crumples and expands like a paper lantern.[239]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e5/Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png/321px-Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png) The mantle of the red paper lantern jellyfish crumples and expands like a paper lantern.[239]
The mantle of the red paper lantern jellyfish crumples and expands like a paper lantern.[239]




![The mantle of the red paper lantern jellyfish crumples and expands like a paper lantern.[239]](/wiki/File:Red-paper-lantern-jellyfish-Karen-Osborn-Smithsonian-Institution.png)
-

-
 Marrus orthocanna another colonial siphonophore, assembled from two types of zooids.
Marrus orthocanna another colonial siphonophore, assembled from two types of zooids. -
![Porpita porpita consists of a colony of hydroids[240]](//upload.wikimedia.org/wikipedia/commons/thumb/2/23/Porpita_porpita.jpg/252px-Porpita_porpita.jpg)
-
![Lion's mane jellyfish, largest known jellyfish[241]](//upload.wikimedia.org/wikipedia/commons/thumb/2/22/Largelionsmanejellyfish.jpg/191px-Largelionsmanejellyfish.jpg)
-
![Turritopsis dohrnii achieves biological immortality by transferring its cells back to childhood.[242][243]](//upload.wikimedia.org/wikipedia/commons/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg) Turritopsis dohrnii achieves biological immortality by transferring its cells back to childhood.[242][243]
Turritopsis dohrnii achieves biological immortality by transferring its cells back to childhood.[242][243] -
![The sea wasp is the most lethal jellyfish in the world.[244]](//upload.wikimedia.org/wikipedia/commons/thumb/7/75/Chironex_fleckeri_%28sea_wasp%29.jpg/255px-Chironex_fleckeri_%28sea_wasp%29.jpg)
.jpg)

![Porpita porpita consists of a colony of hydroids[240]](/wiki/File:Porpita_porpita.jpg)
![Lion's mane jellyfish, largest known jellyfish[241]](/wiki/File:Largelionsmanejellyfish.jpg)
![Turritopsis dohrnii achieves biological immortality by transferring its cells back to childhood.[242][243]](/wiki/File:Turritopsis_dohrnii_(cropped).jpg)
![The sea wasp is the most lethal jellyfish in the world.[244]](/wiki/File:Chironex_fleckeri_(sea_wasp).jpg)
Bilateral invertebrate animals
edit
Some of the earliest bilaterians were wormlike, and the original bilaterian may have been a bottom dwelling worm with a single body opening.[245] A bilaterian body can be conceptualized as a cylinder with a gut running between two openings, the mouth and the anus. Around the gut it has an internal body cavity, a coelom or pseudocoelom.[a] Animals with this bilaterally symmetric body plan have a head (anterior) end and a tail (posterior) end as well as a back (dorsal) and a belly (ventral); therefore they also have a left side and a right side.[246][247]
Having a front end means that this part of the body encounters stimuli, such as food, favouring cephalisation, the development of a head with sense organs and a mouth.[248] The body stretches back from the head, and many bilaterians have a combination of circular muscles that constrict the body, making it longer, and an opposing set of longitudinal muscles, that shorten the body;[247] these enable soft-bodied animals with a hydrostatic skeleton to move by peristalsis.[249] They also have a gut that extends through the basically cylindrical body from mouth to anus. Many bilaterian phyla have primary larvae which swim with cilia and have an apical organ containing sensory cells. However, there are exceptions to each of these characteristics; for example, adult echinoderms are radially symmetric (unlike their larvae), and certain parasitic worms have extremely simplified body structures.[246][247]
.jpg)
| ← bilaterians |
| |||||||||||||||||||||
Protostomes
editProtostomes (from Greek for first mouth) are a superphylum of animals. It is a sister clade of the deuterostomes (from Greek for second mouth), with which it forms the Nephrozoa clade. Protostomes are distinguished from deuterostomes by the way their embryos develop. In protostomes the first opening that develops becomes the mouth, while in deuterostomes it becomes the anus.[251][252]
| ← Protostomes |
| ||||||||||||||||||
| (extant) |


Marine worms
edit
Worms (Old English for serpents) form a number of phyla. Different groups of marine worms are related only distantly, so they are found in several different phyla such as the Annelida (segmented worms), Chaetognatha (arrow worms), Phoronida (horseshoe worms), and Hemichordata. All worms, apart from the Hemichordata, are protostomes. The Hemichordata are deuterostomes and are discussed in their own section below.
The typical body plan of a worm involves long cylindrical tube-like bodies and no limbs. Marine worms vary in size from microscopic to over 1 metre (3.3 ft) in length for some marine polychaete worms (bristle worms)[253] and up to 58 metres (190 ft) for the marine nemertean worm (bootlace worm).[254] Some marine worms occupy a small variety of parasitic niches, living inside the bodies of other animals, while others live more freely in the marine environment or by burrowing underground. Many of these worms have specialized tentacles used for exchanging oxygen and carbon dioxide and also may be used for reproduction. Some marine worms are tube worms, such as the giant tube worm which lives in waters near underwater volcanoes and can withstand temperatures up to 90 degrees Celsius. Platyhelminthes (flatworms) form another worm phylum which includes a class of parasitic tapeworms. The marine tapeworm Polygonoporus giganticus, found in the gut of sperm whales, can grow to over 30 m (100 ft).[255][256]
Nematodes (roundworms) constitute a further worm phylum with tubular digestive systems and an opening at both ends.[257][258] Over 25,000 nematode species have been described,[259][260] of which more than half are parasitic. It has been estimated another million remain undescribed.[261] They are ubiquitous in marine, freshwater and terrestrial environments, where they often outnumber other animals in both individual and species counts. They are found in every part of the Earth's lithosphere, from the top of mountains to the bottom of oceanic trenches.[262] By count they represent 90% of all animals on the ocean floor.[263] Their numerical dominance, often exceeding a million individuals per square meter and accounting for about 80% of all individual animals on Earth, their diversity of life cycles, and their presence at various trophic levels point at an important role in many ecosystems.[264]
-
 Giant tube worms cluster around hydrothermal vents.
Giant tube worms cluster around hydrothermal vents. -
 Nematodes are ubiquitous pseudocoelomates which can parasite marine plants and animals.
Nematodes are ubiquitous pseudocoelomates which can parasite marine plants and animals. -
 Bloodworms are typically found on the bottom of shallow marine waters.
Bloodworms are typically found on the bottom of shallow marine waters.



Marine molluscs
edit



Molluscs (Latin for soft) form a phylum with about 85,000 extant recognized species.[267] They are the largest marine phylum in terms of species count, containing about 23% of all the named marine organisms.[268] Molluscs have more varied forms than other invertebrate phyla. They are highly diverse, not just in size and in anatomical structure, but also in behaviour and in habitat.
.jpg)
The mollusc phylum is divided into 9 or 10 taxonomic classes. These classes include gastropods, bivalves and cephalopods, as well as other lesser-known but distinctive classes. Gastropods with protective shells are referred to as snails, whereas gastropods without protective shells are referred to as slugs. Gastropods are by far the most numerous molluscs in terms of species.[269] Bivalves include clams, oysters, cockles, mussels, scallops, and numerous other families. There are about 8,000 marine bivalves species (including brackish water and estuarine species). A deep sea ocean quahog clam has been reported as having lived 507 years[270] making it the longest recorded life of all animals apart from colonial animals, or near-colonial animals like sponges.[214]
- Gastropods and bivalves
-

-
 The sea snail Syrinx aruanus has a shell up to 91 cm long, the largest of any living gastropod.
The sea snail Syrinx aruanus has a shell up to 91 cm long, the largest of any living gastropod. -
 Molluscs usually have eyes. Bordering the edge of the mantle of a scallop, a bivalve mollusc, can be over 100 simple eyes.
Molluscs usually have eyes. Bordering the edge of the mantle of a scallop, a bivalve mollusc, can be over 100 simple eyes. -
 Common mussel, another bivalve
Common mussel, another bivalve




Cephalopods include octopus, squid and cuttlefish. About 800 living species of marine cephalopods have been identified,[271] and an estimated 11,000 extinct taxa have been described.[272] They are found in all oceans, but there are no fully freshwater cephalopods.[273]
- Cephalopods
-
![The nautilus is a living fossil little changed since it evolved 500 million years ago as one of the first cephalopods.[274][275][276]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/300px-Nautilus_Palau.JPG) The nautilus is a living fossil little changed since it evolved 500 million years ago as one of the first cephalopods.[274][275][276]
The nautilus is a living fossil little changed since it evolved 500 million years ago as one of the first cephalopods.[274][275][276] -

-

-
![Colossal squid, the largest of all invertebrates[277]](//upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/157px-Calmarcolossal.jpg) Colossal squid, the largest of all invertebrates[277]
Colossal squid, the largest of all invertebrates[277]
![The nautilus is a living fossil little changed since it evolved 500 million years ago as one of the first cephalopods.[274][275][276]](/wiki/File:Nautilus_Palau.JPG)


![Colossal squid, the largest of all invertebrates[277]](/wiki/File:Calmarcolossal.jpg)
Molluscs have such diverse shapes that many textbooks base their descriptions of molluscan anatomy on a generalized or hypothetical ancestral mollusc. This generalized mollusc is unsegmented and bilaterally symmetrical with an underside consisting of a single muscular foot. Beyond that it has three further key features. Firstly, it has a muscular cloak called a mantle covering its viscera and containing a significant cavity used for breathing and excretion. A shell secreted by the mantle covers the upper surface. Secondly (apart from bivalves) it has a rasping tongue called a radula used for feeding. Thirdly, it has a nervous system including a complex digestive system using microscopic, muscle-powered hairs called cilia to exude mucus. The generalized mollusc has two paired nerve cords (three in bivalves). The brain, in species that have one, encircles the esophagus. Most molluscs have eyes and all have sensors detecting chemicals, vibrations, and touch.[278][279]
Good evidence exists for the appearance of marine gastropods, cephalopods and bivalves in the Cambrian period 538.8 to 485.4 million years ago.
Marine arthropods
edit

Arthropods (Greek for jointed feet) have an exoskeleton (external skeleton), a segmented body, and jointed appendages (paired appendages). They form a phylum which includes insects, arachnids, myriapods, and crustaceans. Arthropods are characterized by their jointed limbs and cuticle made of chitin, often mineralised with calcium carbonate. The arthropod body plan consists of segments, each with a pair of appendages. The rigid cuticle inhibits growth, so arthropods replace it periodically by moulting. Their versatility has enabled them to become the most species-rich members of all ecological guilds in most environments.
The evolutionary ancestry of arthropods dates back to the Cambrian period and is generally regarded as monophyletic. However, basal relationships of arthropods with extinct phyla such as lobopodians have recently been debated.[282][283]
| Panarthropoda |
| ||||||||||||


- Arthropod fossils and living fossils
-
![Fossil trilobite. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[285]](//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Cheirurus_ingricus.png/301px-Cheirurus_ingricus.png) Fossil trilobite. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[285]
Fossil trilobite. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[285] -
 The Anomalocaris ("abnormal shrimp") was one of the first apex predators and first appeared about 515 Ma.
The Anomalocaris ("abnormal shrimp") was one of the first apex predators and first appeared about 515 Ma. -
![The largest known arthropod, the sea scorpion Jaekelopterus rhenaniae, has been found in estuarine strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[286][287]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/Jaekelopterus_rhenaniae_reconstruction.jpg/331px-Jaekelopterus_rhenaniae_reconstruction.jpg) The largest known arthropod, the sea scorpion Jaekelopterus rhenaniae, has been found in estuarine strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[286][287]
The largest known arthropod, the sea scorpion Jaekelopterus rhenaniae, has been found in estuarine strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[286][287] -
![Xiphosurans, the group including modern Horseshoe crabs appeared around 480 Ma.[288]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d4/Limulus_%28cropped%29.jpg/257px-Limulus_%28cropped%29.jpg)
![Fossil trilobite. Trilobites first appeared about 521 Ma. They were highly successful and were found everywhere in the ocean for 270 Ma.[285]](/wiki/File:Cheirurus_ingricus.png)

![The largest known arthropod, the sea scorpion Jaekelopterus rhenaniae, has been found in estuarine strata from about 390 Ma. It was up to 2.5 m (8.2 ft) long.[286][287]](/wiki/File:Jaekelopterus_rhenaniae_reconstruction.jpg)
![Xiphosurans, the group including modern Horseshoe crabs appeared around 480 Ma.[288]](/wiki/File:Limulus_(cropped).jpg)
Extant marine arthropods range in size from the microscopic crustacean Stygotantulus to the Japanese spider crab. Arthropods' primary internal cavity is a hemocoel, which accommodates their internal organs, and through which their haemolymph - analogue of blood - circulates; they have open circulatory systems. Like their exteriors, the internal organs of arthropods are generally built of repeated segments. Their nervous system is "ladder-like", with paired ventral nerve cords running through all segments and forming paired ganglia in each segment. Their heads are formed by fusion of varying numbers of segments, and their brains are formed by fusion of the ganglia of these segments and encircle the esophagus. The respiratory and excretory systems of arthropods vary, depending as much on their environment as on the subphylum to which they belong.
- Modern crustaceans
-
 Many crustaceans are very small, like this tiny amphipod, and make up a significant part of the ocean's zooplankton.
Many crustaceans are very small, like this tiny amphipod, and make up a significant part of the ocean's zooplankton. -
![The Japanese spider crab has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[289]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a7/Macrocheira_kaempferi.jpg/314px-Macrocheira_kaempferi.jpg) The Japanese spider crab has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[289]
The Japanese spider crab has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[289] -
![The Tasmanian giant crab is long-lived and slow-growing, making it vulnerable to overfishing.[290]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/330px-J_J_Wild_Pseudocarcinus_cropped.jpg) The Tasmanian giant crab is long-lived and slow-growing, making it vulnerable to overfishing.[290]
The Tasmanian giant crab is long-lived and slow-growing, making it vulnerable to overfishing.[290] -
![Mantis shrimp have the most advanced eyes in the animal kingdom,[291] and smash prey by swinging their club-like raptorial claws.[292]](//upload.wikimedia.org/wikipedia/commons/thumb/0/03/Odontodactylus_scyllarus_2.png/298px-Odontodactylus_scyllarus_2.png) Mantis shrimp have the most advanced eyes in the animal kingdom,[291] and smash prey by swinging their club-like raptorial claws.[292]
Mantis shrimp have the most advanced eyes in the animal kingdom,[291] and smash prey by swinging their club-like raptorial claws.[292]

![The Japanese spider crab has the longest leg span of any arthropod, reaching 5.5 metres (18 ft) from claw to claw.[289]](/wiki/File:Macrocheira_kaempferi.jpg)
![The Tasmanian giant crab is long-lived and slow-growing, making it vulnerable to overfishing.[290]](/wiki/File:J_J_Wild_Pseudocarcinus_cropped.jpg)
![Mantis shrimp have the most advanced eyes in the animal kingdom,[291] and smash prey by swinging their club-like raptorial claws.[292]](/wiki/File:Odontodactylus_scyllarus_2.png)
Arthropod vision relies on various combinations of compound eyes and pigment-pit ocelli: in most species the ocelli can only detect the direction from which light is coming, and the compound eyes are the main source of information. Arthropods also have a wide range of chemical and mechanical sensors, mostly based on modifications of the many setae (bristles) that project through their cuticles. Arthropod methods of reproduction are diverse: terrestrial species use some form of internal fertilization while marine species lay eggs using either internal or external fertilization. Arthropod hatchlings vary from miniature adults to grubs that lack jointed limbs and eventually undergo a total metamorphosis to produce the adult form.
Deuterostomes
editIn deuterostomes the first opening that develops in the growing embryo becomes the anus, while in protostomes it becomes the mouth. Deuterostomes form a superphylum of animals and are the sister clade of the protostomes.[251][252] It is once considered that the earliest known deuterostomes are Saccorhytus fossils from about 540 million years ago.[293] However, another study considered that Saccorhytus is more likely to be an ecdysozoan.[294]
| ← deuterostomes | |
| (extant) |

Echinoderms
edit
Echinoderms (Greek for spiny skin) is a phylum which contains only marine invertebrates. The phylum contains about 7000 living species,[295] making it the second-largest grouping of deuterostomes, after the chordates.
Adult echinoderms are recognizable by their radial symmetry (usually five-point) and include starfish, sea urchins, sand dollars, and sea cucumbers, as well as the sea lilies.[296] Echinoderms are found at every ocean depth, from the intertidal zone to the abyssal zone. They are unique among animals in having bilateral symmetry at the larval stage, but fivefold symmetry (pentamerism, a special type of radial symmetry) as adults.[297]
Echinoderms are important both biologically and geologically. Biologically, there are few other groupings so abundant in the biotic desert of the deep sea, as well as shallower oceans. Most echinoderms are able to regenerate tissue, organs, limbs, and reproduce asexually; in some cases, they can undergo complete regeneration from a single limb. Geologically, the value of echinoderms is in their ossified skeletons, which are major contributors to many limestone formations, and can provide valuable clues as to the geological environment. They were the most used species in regenerative research in the 19th and 20th centuries.
-
 Echinoderm literally means "spiny skin", as this water melon sea urchin illustrates.
Echinoderm literally means "spiny skin", as this water melon sea urchin illustrates. -
![The ochre sea star was the first keystone predator to be studied. They limit mussels which can overwhelm intertidal communities.[298]](//upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/360px-Ochre_sea_star.jpg) The ochre sea star was the first keystone predator to be studied. They limit mussels which can overwhelm intertidal communities.[298]
The ochre sea star was the first keystone predator to be studied. They limit mussels which can overwhelm intertidal communities.[298] -
 Colorful sea lilies in shallow waters
Colorful sea lilies in shallow waters

![The ochre sea star was the first keystone predator to be studied. They limit mussels which can overwhelm intertidal communities.[298]](/wiki/File:Ochre_sea_star.jpg)

-
 Sea cucumbers filter feed on plankton and suspended solids.
Sea cucumbers filter feed on plankton and suspended solids. -
 The sea pig, a deep water sea cucumber, is the only echinoderm that uses legged locomotion.
The sea pig, a deep water sea cucumber, is the only echinoderm that uses legged locomotion. -
 A benthopelagic and bioluminescent swimming sea cucumber, 3200 metres deep
A benthopelagic and bioluminescent swimming sea cucumber, 3200 metres deep

.jpg)

It is held by some scientists that the radiation of echinoderms was responsible for the Mesozoic Marine Revolution. Aside from the hard-to-classify Arkarua (a Precambrian animal with echinoderm-like pentamerous radial symmetry), the first definitive members of the phylum appeared near the start of the Cambrian.
Hemichordates
edit

Hemichordates form a sister phylum to the echinoderms. They are solitary worm-shaped organisms rarely seen by humans because of their lifestyle. They include two main groups, the acorn worms and the Pterobranchia. Pterobranchia form a class containing about 30 species of small worm-shaped animals that live in secreted tubes on the ocean floor. Acorn worms form a class containing about 111 species that generally live in U-shaped burrows on the seabed, from the shoreline to a depth of 3000 metres. The worms lie there with the proboscis sticking out of one opening in the burrow, subsisting as deposit feeders or suspension feeders. It is supposed the ancestors of acorn worms used to live in tubes like their relatives, the Pterobranchia, but eventually started to live a safer and more sheltered existence in sediment burrows.[302] Some of these worms may grow to be very long; one particular species may reach a length of 2.5 metres (8 ft 2 in), although most acorn worms are much smaller.
Acorn worms are more highly specialised and advanced than other worm-like organisms. They have a circulatory system with a heart that also functions as a kidney. Acorn worms have gill-like structures they use for breathing, similar to the gills of fish. Therefore, acorn worms are sometimes said to be a link between classical invertebrates and vertebrates. Acorn worms continually form new gill slits as they grow in size, and some older individuals have more than a hundred on each side. Each slit consists of a branchial chamber opening to the pharynx through a U-shaped cleft. Cilia push water through the slits, maintaining a constant flow, just as in fish.[303] Some acorn worms also have a postanal tail which may be homologous to the post-anal tail of vertebrates.
The three-section body plan of the acorn worm is no longer present in the vertebrates, except in the anatomy of the frontal neural tube, later developed into a brain divided into three parts. This means some of the original anatomy of the early chordate ancestors is still present in vertebrates even if it is not always visible. One theory is the three-part body originated from an early common ancestor of the deuterostomes, and maybe even from a common bilateral ancestor of both deuterostomes and protostomes. Studies have shown the gene expression in the embryo share three of the same signaling centers that shape the brains of all vertebrates, but instead of taking part in the formation of their neural system,[304] they are controlling the development of the different body regions.[305]
Marine chordates
edit
The chordate phylum has three subphyla, one of which is the vertebrates (see below). The other two subphyla are marine invertebrates: the tunicates (salps and sea squirts) and the cephalochordates (such as lancelets). Invertebrate chordates are close relatives to vertebrates. In particular, there has been discussion about how closely some extinct marine species, such as Pikaiidae, Palaeospondylus, Zhongxiniscus and Vetulicolia, might relate ancestrally to vertebrates.
- Invertebrate chordates are close relatives of vertebrates
-
![The lancelet, a small translucent fish-like cephalochordate, is one of the closest living invertebrate relative of the vertebrates.[307][308]](//upload.wikimedia.org/wikipedia/commons/thumb/5/54/Amphioxus.png/332px-Amphioxus.png) The lancelet, a small translucent fish-like cephalochordate, is one of the closest living invertebrate relative of the vertebrates.[307][308]
The lancelet, a small translucent fish-like cephalochordate, is one of the closest living invertebrate relative of the vertebrates.[307][308] -
![Tunicates, like these fluorescent-colored sea squirts, may provide clues to vertebrate and therefore human ancestry.[309]](//upload.wikimedia.org/wikipedia/commons/thumb/8/8c/Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png/280px-Ascidian_%28Rhopalaea_Crassa%29_%284_cm%29.png) Tunicates, like these fluorescent-colored sea squirts, may provide clues to vertebrate and therefore human ancestry.[309]
Tunicates, like these fluorescent-colored sea squirts, may provide clues to vertebrate and therefore human ancestry.[309] -
 Pyrosomes are free-floating bioluminescent tunicates made up of hundreds of individuals.
Pyrosomes are free-floating bioluminescent tunicates made up of hundreds of individuals. -
 Salp chain
Salp chain
![The lancelet, a small translucent fish-like cephalochordate, is one of the closest living invertebrate relative of the vertebrates.[307][308]](/wiki/File:Amphioxus.png)
![Tunicates, like these fluorescent-colored sea squirts, may provide clues to vertebrate and therefore human ancestry.[309]](/wiki/File:Ascidian_(Rhopalaea_Crassa)_(4_cm).png)

.jpg)



Vertebrate animals
edit

Vertebrates (Latin for joints of the spine) are a subphylum of chordates. They are chordates that have a vertebral column (backbone). The vertebral column provides the central support structure for an internal skeleton which gives shape, support, and protection to the body and can provide a means of anchoring fins or limbs to the body. The vertebral column also serves to house and protect the spinal cord that lies within the vertebral column.
Marine vertebrates can be divided into marine fish and marine tetrapods.
Marine fish
editFish typically breathe by extracting oxygen from water through gills and have a skin protected by scales and mucous. They use fins to propel and stabilise themselves in the water, and usually have a two-chambered heart and eyes well adapted to seeing underwater, as well as other sensory systems. Over 33,000 species of fish have been described as of 2017,[310] of which about 20,000 are marine fish.[311]
| ← vertebrates | |
| (extant) |
Jawless fish
edit
Early fish had no jaws. Most went extinct when they were outcompeted by jawed fish (below), but two groups survived: hagfish and lampreys. Hagfish form a class of about 20 species of eel-shaped, slime-producing marine fish. They are the only known living animals that have a skull but no vertebral column. Lampreys form a superclass containing 38 known extant species of jawless fish.[312] The adult lamprey is characterized by a toothed, funnel-like sucking mouth. Although they are well known for boring into the flesh of other fish to suck their blood,[313] only 18 species of lampreys are actually parasitic.[314] Together hagfish and lampreys are the sister group to vertebrates. Living hagfish remain similar to hagfish from around 300 million years ago.[315] The lampreys are a very ancient lineage of vertebrates, though their exact relationship to hagfishes and jawed vertebrates is still a matter of dispute.[316] Molecular analysis since 1992 has suggested that hagfish are most closely related to lampreys,[317] and so also are vertebrates in a monophyletic sense. Others consider them a sister group of vertebrates in the common taxon of craniata.[318]
The Tully monster is an extinct genus of soft-bodied bilaterians that lived in tropical estuaries about 300 million years ago. Since 2016 there has been controversy over whether this animal was a vertebrate or an invertebrate.[319][320] In 2020 researchers found "strong evidence" that the Tully monster was a vertebrate, and was a jawless fish in the lineage of the lamprey,[321][322] while in 2023 other researchers found 3D fossils scans did not support those conclusions.[323]
-

-
 Lampreys are often parasitic and have a toothed, funnel-like sucking mouth.
Lampreys are often parasitic and have a toothed, funnel-like sucking mouth. -
 The extinct Pteraspidomorphi, ancestral to jawed vertebrates
The extinct Pteraspidomorphi, ancestral to jawed vertebrates



Pteraspidomorphi is an extinct class of early jawless fish ancestral to jawed vertebrates. The few characteristics they share with the latter are now considered as primitive for all vertebrates.
Around the start of the Devonian, fish started appearing with a deep remodelling of the vertebrate skull that resulted in a jaw.[324] All vertebrate jaws, including the human jaw, have evolved from these early fish jaws. The appearance of the early vertebrate jaw has been described as "perhaps the most profound and radical evolutionary step in vertebrate history".[325][326] Jaws make it possible to capture, hold, and chew prey. Fish without jaws had more difficulty surviving than fish with jaws, and most jawless fish became extinct during the Triassic period.
Cartilaginous fish
editJawed fish fall into two main groups: fish with bony internal skeletons and fish with cartilaginous internal skeletons. Cartilaginous fish, such as sharks and rays, have jaws and skeletons made of cartilage rather than bone. Megalodon is an extinct species of shark that lived about 28 to 1.5 Ma. It may looked much like a stocky version of the great white shark, but was much larger with estimated lengths reaching 20.3 metres (67 ft).[327] Found in all oceans[328] it was one of the largest and most powerful predators in vertebrate history,[327] and probably had a profound impact on marine life.[329] The Greenland shark has the longest known lifespan of all vertebrates, about 400 years.[330] Some sharks such as the great white are partially warm blooded and give live birth. The manta ray, largest ray in the world, has been targeted by fisheries and is now vulnerable.[331]
- Cartilaginous fishes
-
 Cartilaginous fishes may have evolved from spiny sharks.
Cartilaginous fishes may have evolved from spiny sharks. -

-
 Manta ray, the largest ray
Manta ray, the largest ray -
![Sawfish, rays with long rostrums resembling a saw. All species are now endangered.[332]](//upload.wikimedia.org/wikipedia/commons/thumb/4/48/Pristis_clavata_2.jpg/407px-Pristis_clavata_2.jpg)



![Sawfish, rays with long rostrums resembling a saw. All species are now endangered.[332]](/wiki/File:Pristis_clavata_2.jpg)
-
 The extinct megalodon resembled a giant great white shark.
The extinct megalodon resembled a giant great white shark. -
 The Greenland shark lives longer than any other vertebrate.
The Greenland shark lives longer than any other vertebrate. -



.jpg)
Bony fish
edit

.jpg)
Bony fish have jaws and skeletons made of bone rather than cartilage. Bony fish also have hard, bony plates called operculum which help them respire and protect their gills, and they often possess a swim bladder which they use for better control of their buoyancy. Bony fish can be further divided into those with lobe fins and those with ray fins. The approximate dates in the phylogenetic tree are from Near et al., 2012[333] and Zhu et al., 2009.[334]
| ← bony fish |
| ||||||||||||||||||
| (extant) |
Lobe fins have the form of fleshy lobes supported by bony stalks which extend from the body.[335] Guiyu oneiros, the earliest-known bony fish, lived during the Late Silurian 419 million years ago. It has the combination of both ray-finned and lobe-finned features, although analysis of the totality of its features place it closer to lobe-finned fish.[334] Lobe fins evolved into the legs of the first tetrapod land vertebrates, so by extension an early ancestor of humans was a lobe-finned fish. Apart from the coelacanths and the lungfishes, lobe-finned fishes are now extinct.
The remaining bony fish have ray fins. These are made of webs of skin supported by bony or horny spines (rays) which can be erected to control the fin stiffness.
- The main distinguishing feature of the chondrosteans (sturgeon, paddlefish, bichir and reedfish) is the cartilaginous nature of their skeletons. The ancestors of the chondrosteans are thought to be bony fish, but the characteristic of an ossified skeleton was lost in later evolutionary development, resulting in a lightening of the frame.[336]
- Neopterygians (from Greek for new fins) appeared sometime in the Late Permian, before dinosaurs. They were a very successful group of fish, because they could move more rapidly than their ancestors. Their scales and skeletons began to lighten during their evolution, and their jaws became more powerful and efficient.[337]
Teleosts
edit
About 96% of all modern fish species are teleosts,[338] of which about 14,000 are marine species.[339] Teleosts can be distinguished from other bony fish by their possession of a homocercal tail, a tail where the upper half mirrors the lower half.[340] Another difference lies in their jaw bones – teleosts have modifications in the jaw musculature which make it possible for them to protrude their jaws. This enables them to grab prey and draw it into their mouth.[340] In general, teleosts tend to be quicker and more flexible than more basal bony fishes. Their skeletal structure has evolved towards greater lightness. While teleost bones are well calcified, they are constructed from a scaffolding of struts, rather than the dense cancellous bones of holostean fish.[341]
Teleosts are found in almost all marine habitats.[342] They have enormous diversity, and range in size from adult gobies 8mm long [343] to ocean sunfish weighing over 2,000 kg.[344] The following images show something of the diversity in the shape and colour of modern marine teleosts...








Nearly half of all extant vertebrate species are teleosts.[345]
Marine tetrapods
edit
A tetrapod (Greek for four feet) is a vertebrate with limbs (feet). Tetrapods evolved from ancient lobe-finned fishes about 400 million years ago during the Devonian Period when their earliest ancestors emerged from the sea and adapted to living on land.[346] This change from a body plan for breathing and navigating in gravity-neutral water to a body plan with mechanisms enabling the animal to breath in air without dehydrating and move on land is one of the most profound evolutionary changes known.[347][348] Tetrapods can be divided into four classes: amphibians, reptiles, birds and mammals.
| ← tetrapods |
| ||||||||||||||||||
Marine tetrapods are tetrapods that returned from land back to the sea again. The first returns to the ocean may have occurred as early as the Carboniferous Period[349] whereas other returns occurred as recently as the Cenozoic, as in cetaceans, pinnipeds,[350] and several modern amphibians.[351] Amphibians (from Greek for both kinds of life) live part of their life in water and part on land. They mostly require fresh water to reproduce. A few inhabit brackish water, but there are no true marine amphibians.[352] There have been reports, however, of amphibians invading marine waters, such as a Black Sea invasion by the natural hybrid Pelophylax esculentus reported in 2010.[353]
Reptiles
editReptiles (Late Latin for creeping or crawling) do not have an aquatic larval stage, and in this way are unlike amphibians. Most reptiles are oviparous, although several species of squamates are viviparous, as were some extinct aquatic clades[354] — the fetus develops within the mother, contained in a placenta rather than an eggshell. As amniotes, reptile eggs are surrounded by membranes for protection and transport, which adapt them to reproduction on dry land. Many of the viviparous species feed their fetuses through various forms of placenta analogous to those of mammals, with some providing initial care for their hatchlings.
Some reptiles are more closely related to birds than other reptiles, and many scientists prefer to make Reptilia a monophyletic group which includes the birds.[355][356][357][358] Extant non-avian reptiles which inhabit or frequent the sea include sea turtles, sea snakes, terrapins, the marine iguana, and the saltwater crocodile. Currently, of the approximately 12,000 extant reptile species and sub-species, only about 100 of are classed as marine reptiles.[359]
Except for some sea snakes, most extant marine reptiles are oviparous and need to return to land to lay their eggs. Apart from sea turtles, the species usually spend most of their lives on or near land rather than in the ocean. Sea snakes generally prefer shallow waters nearby land, around islands, especially waters that are somewhat sheltered, as well as near estuaries.[360][361] Unlike land snakes, sea snakes have evolved flattened tails which help them swim.[362]
-

-

-

-
 Marine snakes have flattened tails.
Marine snakes have flattened tails. -
 The ancient Ichthyosaurus communis independently evolved flippers similar to dolphins.
The ancient Ichthyosaurus communis independently evolved flippers similar to dolphins.

.jpg)
.jpg)


Some extinct marine reptiles, such as ichthyosaurs, evolved to be viviparous and had no requirement to return to land. Ichthyosaurs resembled dolphins. They first appeared about 245 million years ago and disappeared about 90 million years ago. The terrestrial ancestor of the ichthyosaur had no features already on its back or tail that might have helped along the evolutionary process. Yet the ichthyosaur developed a dorsal and tail fin which improved its ability to swim.[363] The biologist Stephen Jay Gould said the ichthyosaur was his favourite example of convergent evolution.[364] The earliest marine reptiles arose in the Permian. During the Mesozoic many groups of reptiles became adapted to life in the seas, including ichthyosaurs, plesiosaurs, mosasaurs, nothosaurs, placodonts, sea turtles, thalattosaurs and thalattosuchians. Marine reptiles were less numerous after mass extinction at the end of the Cretaceous.
Birds
edit
Marine birds are adapted to life within the marine environment. They are often called seabirds. While marine birds vary greatly in lifestyle, behaviour and physiology, they often exhibit striking convergent evolution, as the same environmental problems and feeding niches have resulted in similar adaptations. Examples include albatross, penguins, gannets, and auks.
In general, marine birds live longer, breed later and have fewer young than terrestrial birds do, but they invest a great deal of time in their young. Most species nest in colonies, which can vary in size from a few dozen birds to millions. Many species are famous for undertaking long annual migrations, crossing the equator or circumnavigating the Earth in some cases. They feed both at the ocean's surface and below it, and even feed on each other. Marine birds can be highly pelagic, coastal, or in some cases spend a part of the year away from the sea entirely. Some marine birds plummet from heights, plunging through the water leaving vapour-like trails, similar to that of fighter planes.[365] Gannets plunge into the water at up to 100 kilometres per hour (60 mph). They have air sacs under their skin in their face and chest which act like bubble-wrap, cushioning the impact with the water.
-
 European herring gull attack herring schools from above.
European herring gull attack herring schools from above. -
 Gentoo penguin swimming underwater
Gentoo penguin swimming underwater -
 Albatrosses range over huge areas of ocean and some even circle the globe.
Albatrosses range over huge areas of ocean and some even circle the globe. -
 Gannets "divebomb" at high speed
Gannets "divebomb" at high speed




The first marine birds evolved in the Cretaceous period, and modern marine bird families emerged in the Paleogene.
Mammals
edit
Mammals (from Latin for breast) are characterised by the presence of mammary glands which in females produce milk for feeding (nursing) their young. There are about 130 living and recently extinct marine mammal species such as seals, dolphins, whales, manatees, sea otters and polar bears.[366] They do not represent a distinct taxon or systematic grouping, but are instead unified by their reliance on the marine environment for feeding. Both cetaceans and sirenians are fully aquatic and therefore are obligate water dwellers. Seals and sea-lions are semiaquatic; they spend the majority of their time in the water, but need to return to land for important activities such as mating, breeding and molting. In contrast, both otters and the polar bear are much less adapted to aquatic living. Their diet varies considerably as well: some may eat zooplankton; others may eat fish, squid, shellfish, and sea-grass; and a few may eat other mammals.
In a process of convergent evolution, marine mammals, especially cetaceans such as dolphins and whales, redeveloped their body plan to parallel the streamlined fusiform body plan of pelagic fish. Front legs became flippers and back legs disappeared, a dorsal fin reappeared and the tail morphed into a powerful horizontal fluke. This body plan is an adaptation to being an active predator in a high drag environment. A parallel convergence occurred with the now extinct marine reptile ichthyosaur.[367]
-
![Endangered blue whale, the largest living animal[368]](//upload.wikimedia.org/wikipedia/commons/thumb/c/cd/Bluewhale2_noaa.jpg/270px-Bluewhale2_noaa.jpg) Endangered blue whale, the largest living animal[368]
Endangered blue whale, the largest living animal[368] -
![The bottlenose dolphin has the highest encephalization of any animal after humans[369]](//upload.wikimedia.org/wikipedia/commons/thumb/1/10/Tursiops_truncatus_01.jpg/271px-Tursiops_truncatus_01.jpg)
-

-

-

-

![Endangered blue whale, the largest living animal[368]](/wiki/File:Bluewhale2_noaa.jpg)
![The bottlenose dolphin has the highest encephalization of any animal after humans[369]](/wiki/File:Tursiops_truncatus_01.jpg)



.jpg)
Primary producers
edit
Primary producers are the autotroph organisms that make their own food instead of eating other organisms. This means primary producers become the starting point in the food chain for heterotroph organisms that do eat other organisms. Some marine primary producers are specialised bacteria and archaea which are chemotrophs, making their own food by gathering around hydrothermal vents and cold seeps and using chemosynthesis. However most marine primary production comes from organisms which use photosynthesis on the carbon dioxide dissolved in the water. This process uses energy from sunlight to convert water and carbon dioxide[370]: 186–187 into sugars that can be used both as a source of chemical energy and of organic molecules that are used in the structural components of cells.[370]: 1242 Marine primary producers are important because they underpin almost all marine animal life by generating most of the oxygen and food that provide other organisms with the chemical energy they need to exist.
The principal marine primary producers are cyanobacteria, algae and marine plants. The oxygen released as a by-product of photosynthesis is needed by nearly all living things to carry out cellular respiration. In addition, primary producers are influential in the global carbon and water cycles. They stabilize coastal areas and can provide habitats for marine animals. The term division has been traditionally used instead of phylum when discussing primary producers, but the International Code of Nomenclature for algae, fungi, and plants now accepts both terms as equivalents.[371]
Cyanobacteria
edit
.jpg)
Cyanobacteria were the first organisms to evolve an ability to turn sunlight into chemical energy. They form a phylum (division) of bacteria which range from unicellular to filamentous and include colonial species. They are found almost everywhere on earth: in damp soil, in both freshwater and marine environments, and even on Antarctic rocks.[372] In particular, some species occur as drifting cells floating in the ocean, and as such were amongst the first of the phytoplankton.
The first primary producers that used photosynthesis were oceanic cyanobacteria about 2.3 billion years ago.[373][374] The release of molecular oxygen by cyanobacteria as a by-product of photosynthesis induced global changes in the Earth's environment. Because oxygen was toxic to most life on Earth at the time, this led to the near-extinction of oxygen-intolerant organisms, a dramatic change which redirected the evolution of the major animal and plant species.[375]
The tiny marine cyanobacterium Prochlorococcus, discovered in 1986, forms today part of the base of the ocean food chain and accounts for much of the photosynthesis of the open ocean[376] and an estimated 20% of the oxygen in the Earth's atmosphere.[377] It is possibly the most plentiful genus on Earth: a single millilitre of surface seawater may contain 100,000 cells or more.[378]
Originally, biologists classified cyanobacteria as algae, and referred to it as "blue-green algae". The more recent view is that cyanobacteria are bacteria, and hence are not even in the same Kingdom as algae. Most authorities today exclude all prokaryotes, and hence cyanobacteria from the definition of algae.[379][380]
Algae
edit.jpg)
.jpg)


Algae is an informal term for a widespread and diverse group of photosynthetic protists which are not necessarily closely related and are thus polyphyletic. Marine algae can be divided into six groups:
- green algae, an informal group containing about 8,000 recognised species.[381] Many species live most of their lives as single cells or are filamentous, while others form colonies made up from long chains of cells, or are highly differentiated macroscopic seaweeds.
- red algae, a (disputed) phylum containing about 7,000 recognised species,[382] mostly multicellular and including many notable seaweeds.[382][383]
- brown algae, a class containing about 2,000 recognised species,[384] mostly multicellular and including many seaweeds, including kelp
- diatoms, a (disputed) phylum containing about 100,000 recognised species of mainly unicellular algae. Diatoms generate about 20 percent of the oxygen produced on the planet each year,[147] take in over 6.7 billion metric tons of silicon each year from the waters in which they live,[385] and contribute nearly half of the organic material found in the oceans. The shells (frustules) of dead diatoms can reach as much as half a mile deep on the ocean floor.[386]
- dinoflagellates, a phylum of unicellular flagellates with about 2,000 marine species.[387] Many dinoflagellates are known to be photosynthetic, but a large fraction of these are in fact mixotrophic, combining photosynthesis with ingestion of prey (phagotrophy).[388] Some species are endosymbionts of marine animals and play an important part in the biology of coral reefs. Others predate other protozoa, and a few forms are parasitic.
- euglenophytes, a phylum of unicellular flagellates with only a few marine members
Unlike higher plants, algae lack roots, stems, or leaves. They can be classified by size as microalgae or macroalgae.
Microalgae are the microscopic types of algae, not visible to the naked eye. They are mostly unicellular species which exist as individuals or in chains or groups, though some are multicellular. Microalgae are important components of the marine protists (discussed above), as well as the phytoplankton (discussed below). They are very diverse. It has been estimated there are 200,000-800,000 species of which about 50,000 species have been described.[389] Depending on the species, their sizes range from a few micrometers (μm) to a few hundred micrometers. They are specially adapted to an environment dominated by viscous forces.
-
 Chlamydomonas globosa, a unicellular green alga with two flagella just visible at bottom left
Chlamydomonas globosa, a unicellular green alga with two flagella just visible at bottom left -
![Chlorella vulgaris, a common green microalgae, in endosymbiosis with a ciliate[390]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b6/%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg/293px-%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)
-
 Centric diatom
Centric diatom -
 Dinoflagellates
Dinoflagellates
.jpg)
![Chlorella vulgaris, a common green microalgae, in endosymbiosis with a ciliate[390]](/wiki/File:%D0%98%D0%BD%D1%84%D1%83%D0%B7%D0%BE%D1%80%D0%B8%D0%B8_Ophridium_versatile.jpg)


Macroalgae are the larger, multicellular and more visible types of algae, commonly called seaweeds. Seaweeds usually grow in shallow coastal waters where they are anchored to the seafloor by a holdfast. Seaweed that becomes adrift can wash up on beaches. Kelp is a large brown seaweed that forms large underwater forests covering about 25% of the world coastlines.[391] They are among the most productive and dynamic ecosystems on Earth.[392] Some Sargassum seaweeds are planktonic (free-floating). Like microalgae, macroalgae (seaweeds) are technically marine protists since they are not true plants.
-

-
 Sargassum seaweed is a planktonic brown alga with air bladders that help it float.
Sargassum seaweed is a planktonic brown alga with air bladders that help it float. -
 Sargassum fish are camouflaged to live among drifting Sargassum seaweed.
Sargassum fish are camouflaged to live among drifting Sargassum seaweed.




- Unicellular macroalgae (see also macroscopic protists ←)
-
![The unicellular bubble algae lives in tidal zones. It can have a 4 cm diameter.[393]](//upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Ventricaria_ventricosa.JPG/248px-Ventricaria_ventricosa.JPG)
-
 The unicellular mermaid's wineglass are mushroom-shaped algae that grow up to 10 cm high.
The unicellular mermaid's wineglass are mushroom-shaped algae that grow up to 10 cm high. -
![Killer algae are single-celled organisms, but look like ferns and grow stalks up to 80 cm long.[394]](//upload.wikimedia.org/wikipedia/commons/thumb/e/e7/CaulerpaTaxifolia.jpg/316px-CaulerpaTaxifolia.jpg) Killer algae are single-celled organisms, but look like ferns and grow stalks up to 80 cm long.[394]
Killer algae are single-celled organisms, but look like ferns and grow stalks up to 80 cm long.[394]
![The unicellular bubble algae lives in tidal zones. It can have a 4 cm diameter.[393]](/wiki/File:Ventricaria_ventricosa.JPG)

![Killer algae are single-celled organisms, but look like ferns and grow stalks up to 80 cm long.[394]](/wiki/File:CaulerpaTaxifolia.jpg)
Unicellular organisms are usually microscopic, less than one tenth of a millimeter long. There are exceptions. Mermaid's wineglass, a genus of subtropical green algae, is single-celled but remarkably large and complex in form with a single large nucleus, making it a model organism for studying cell biology.[395] Another single celled algae, Caulerpa taxifolia, has the appearance of a vascular plant including "leaves" arranged neatly up stalks like a fern. Selective breeding in aquariums to produce hardier strains resulted in an accidental release into the Mediterranean where it has become an invasive species known colloquially as killer algae.[396]
Origin of plants
edit
Back in the Silurian, some phytoplankton evolved into red, brown and green algae. These algae then invaded the land and started evolving into the land plants we know today. Later, in the Cretaceous, some of these land plants returned to the sea as marine plants, such as mangroves and seagrasses.[397]
Marine plants can be found in intertidal zones and shallow waters, such as seagrasses like eelgrass and turtle grass, Thalassia. These plants have adapted to the high salinity of the ocean environment. Plant life can also flourish in the brackish waters of estuaries, where mangroves or cordgrass or beach grass beach grass might grow.
-

-

-
![Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows.[398]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Leafy_Sea_Dragon_SA.jpg/120px-Leafy_Sea_Dragon_SA.jpg) Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows.[398]
Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows.[398]


![Sea dragons camouflaged to look like floating seaweed live in kelp forests and seagrass meadows.[398]](/wiki/File:Leafy_Sea_Dragon_SA.jpg)
The total world area of mangrove forests was estimated in 2010 as 134,257 square kilometres (51,837 sq mi) (based on satellite data).[399][400] The total world area of seagrass meadows is more difficult to determine, but was conservatively estimated in 2003 as 177,000 square kilometres (68,000 sq mi).[401]
Mangroves and seagrasses provide important nursery habitats for marine life, acting as hiding and foraging places for larval and juvenile forms of larger fish and invertebrates.[402]
Plankton and trophic interactions
edit
Plankton (from Greek for wanderers) are a diverse group of organisms that live in the water column of large bodies of water but cannot swim against a current. As a result, they wander or drift with the currents.[403] Plankton are defined by their ecological niche, not by any phylogenetic or taxonomic classification. They are a crucial source of food for many marine animals, from forage fish to whales. Plankton can be divided into a plant-like component and an animal component.
Phytoplankton
editPhytoplankton are the plant-like components of the plankton community ("phyto" comes from the Greek for plant). They are autotrophic (self-feeding), meaning they generate their own food and do not need to consume other organisms.
Phytoplankton consist mainly of microscopic photosynthetic eukaryotes which inhabit the upper sunlit layer in all oceans. They need sunlight so they can photosynthesize. Most phytoplankton are single-celled algae, but other phytoplankton are bacteria and some are protists.[404] Phytoplankton groups include cyanobacteria (above), diatoms, various other types of algae (red, green, brown, and yellow-green), dinoflagellates, euglenoids, coccolithophorids, cryptomonads, chrysophytes, chlorophytes, prasinophytes, and silicoflagellates. They form the base of the primary production that drives the ocean food web, and account for half of the current global primary production, more than the terrestrial forests.[405]


- Phytoplankton
-
 Phytoplankton are the foundation of the ocean food chain.
Phytoplankton are the foundation of the ocean food chain. -
 Phytoplankton come in many shapes and sizes.
Phytoplankton come in many shapes and sizes. -
 Diatoms are one of the most common types of phytoplankton.
Diatoms are one of the most common types of phytoplankton. -
 Colonial phytoplankton
Colonial phytoplankton


_Various_diatoms.jpg)

-
 The cyanobacterium Prochlorococcus accounts for much of the ocean's primary production.
The cyanobacterium Prochlorococcus accounts for much of the ocean's primary production. -
 Green cyanobacteria scum washed up on a rock in California
Green cyanobacteria scum washed up on a rock in California -
 Gyrodinium, one of the few naked dinoflagellates which lack armour
Gyrodinium, one of the few naked dinoflagellates which lack armour -
 Zoochlorellae (green) living inside the ciliate Stichotricha secunda
Zoochlorellae (green) living inside the ciliate Stichotricha secunda



.jpg)

-
 Coccolithophores named after the BBC documentary series. The Blue Planet
Coccolithophores named after the BBC documentary series. The Blue Planet -

-
 Algae bloom of Emiliania huxleyi off the southern coast of England
Algae bloom of Emiliania huxleyi off the southern coast of England -
![Guinardia delicatula, a diatom responsible for algal blooms in the North Sea and the English Channel[407]](//upload.wikimedia.org/wikipedia/commons/thumb/4/43/Fjouenne_sbrmvr012w_20070924163039_small.jpg/260px-Fjouenne_sbrmvr012w_20070924163039_small.jpg) Guinardia delicatula, a diatom responsible for algal blooms in the North Sea and the English Channel[407]
Guinardia delicatula, a diatom responsible for algal blooms in the North Sea and the English Channel[407]



![Guinardia delicatula, a diatom responsible for algal blooms in the North Sea and the English Channel[407]](/wiki/File:Fjouenne_sbrmvr012w_20070924163039_small.jpg)
Zooplankton
edit

Zooplankton are the animal component of the planktonic community ("zoo" comes from the Greek for animal). They are heterotrophic (other-feeding), meaning they cannot produce their own food and must consume instead other plants or animals as food. In particular, this means they eat phytoplankton.



.jpg)
Zooplankton are generally larger than phytoplankton, mostly still microscopic but some can be seen with the naked eye. Many protozoans (single-celled protists that prey on other microscopic life) are zooplankton, including zooflagellates, foraminiferans, radiolarians and some dinoflagellates. Other dinoflagellates are mixotrophic and could also be classified as phytoplankton; the distinction between plants and animals often breaks down in very small organisms. Other zooplankton include pelagic cnidarians, ctenophores, molluscs, arthropods and tunicates, as well as planktonic arrow worms and bristle worms.
Radiolarians are unicellular protists with elaborate silica shells
Microzooplankton: major grazers of the plankton
-
 Radiolarians come in many shapes.
Radiolarians come in many shapes. -
 Group of planktic foraminiferans
Group of planktic foraminiferans -
 Copepods eat phytoplankton. This one is carrying eggs.
Copepods eat phytoplankton. This one is carrying eggs. -
 The dinoflagellate, Protoperidinium extrudes a large feeding veil to capture prey.
The dinoflagellate, Protoperidinium extrudes a large feeding veil to capture prey.




Larger zooplankton can be predatory on smaller zooplankton.
Macrozooplankton
-

-

-

-
![Tomopteris, a planktonic segmented worm with unusual yellow bioluminescence[409]](//upload.wikimedia.org/wikipedia/commons/thumb/4/40/Tomopteriskils.jpg/318px-Tomopteriskils.jpg)
-
 Marine amphipod
Marine amphipod -

-
 Pelagic sea cucumber
Pelagic sea cucumber
_Luc_Viatour.jpg)


![Tomopteris, a planktonic segmented worm with unusual yellow bioluminescence[409]](/wiki/File:Tomopteriskils.jpg)

.jpg)

| External videos | |
|---|---|
Many marine animals begin life as zooplankton in the form of eggs or larvae, before they develop into adults. These are meroplanktic, that is, they are planktonic for only part of their life.
- Larvae and juveniles
-
 Salmon larva hatching from its egg
Salmon larva hatching from its egg -
 Ocean sunfish larva
Ocean sunfish larva -
 Juvenile planktonic squid
Juvenile planktonic squid -
 Larva stage of a spiny lobster
Larva stage of a spiny lobster




Mixotrophic plankton
edit
.jpg)
Dinoflagellates are often mixotrophic or live in symbiosis with other organisms.
- Mixoplankton
-
 Tintinnid ciliate Favella
Tintinnid ciliate Favella -
 Euglena mutabilis, a photosynthetic flagellate
Euglena mutabilis, a photosynthetic flagellate -
 Noctiluca scintillans, a bioluminescence dinoflagellate
Noctiluca scintillans, a bioluminescence dinoflagellate


Some dinoflagellates are bioluminescent. At night, ocean water can light up internally and sparkle with blue light because of these dinoflagellates.[411][412] Bioluminescent dinoflagellates possess scintillons, individual cytoplasmic bodies which contain dinoflagellate luciferase, the main enzyme involved in the luminescence. The luminescence, sometimes called the phosphorescence of the sea, occurs as brief (0.1 sec) blue flashes or sparks when individual scintillons are stimulated, usually by mechanical disturbances from, for example, a boat or a swimmer or surf.[413]
Marine food web
edit
Compared to terrestrial environments, marine environments have biomass pyramids which are inverted at the base. In particular, the biomass of consumers (copepods, krill, shrimp, forage fish) is larger than the biomass of primary producers. This happens because the ocean's primary producers are tiny phytoplankton which tend to be r-strategists that grow and reproduce rapidly, so a small mass can have a fast rate of primary production. In contrast, terrestrial primary producers, such as mature forests, are often K-strategists that grow and reproduce slowly, so a much larger mass is needed to achieve the same rate of primary production.
Because of this inversion, it is the zooplankton that make up most of the marine animal biomass. As primary consumers, they are the crucial link between the primary producers (mainly phytoplankton) and the rest of the marine food web (secondary consumers).[414]
If phytoplankton dies before it is eaten, it descends through the euphotic zone as part of the marine snow and settles into the depths of sea. In this way, phytoplankton sequester about 2 billion tons of carbon dioxide into the ocean each year, causing the ocean to become a sink of carbon dioxide holding about 90% of all sequestered carbon.[415]
In 2010 researchers found whales carry nutrients from the depths of the ocean back to the surface using a process they called the whale pump.[416] Whales feed at deeper levels in the ocean where krill is found, but return regularly to the surface to breathe. There whales defecate a liquid rich in nitrogen and iron. Instead of sinking, the liquid stays at the surface where phytoplankton consume it. In the Gulf of Maine the whale pump provides more nitrogen than the rivers.[417]
Other interactions
editBiogeochemical cycles
edit
Taken as a whole, the oceans form a single marine system where water – the "universal solvent" [418] – dissolves nutrients and substances containing elements such as oxygen, carbon, nitrogen and phosphorus. These substances are endlessly cycled and recycled, chemically combined and then broken down again, dissolved and then precipitated or evaporated, imported from and exported back to the land and the atmosphere and the ocean floor. Powered both by the biological activity of marine organisms and by the natural actions of the sun and tides and movements within the Earth's crust, these are the marine biogeochemical cycles.[419][420]
![Marine carbon cycle[421]](/wiki/File:OceanCarbonCycle.jpg)

-
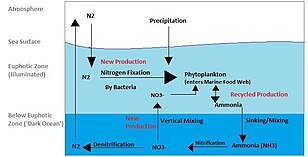
-
 Marine phosphorus cycle
Marine phosphorus cycle


Sediments and biogenic ooze
edit.jpg)
Sediments at the bottom of the ocean have two main origins, terrigenous and biogenous. Terrigenous sediments account for about 45% of the total marine sediment, and originate in the erosion of rocks on land, transported by rivers and land runoff, windborne dust, volcanoes, or grinding by glaciers.
Biogenous sediments account for the other 55% of the total sediment, and originate in the skeletal remains of marine protists (single-celled plankton and benthos organisms). Much smaller amounts of precipitated minerals and meteoric dust can also be present. Ooze, in the context of a marine sediment, does not refer to the consistency of the sediment but to its biological origin. The term ooze was originally used by John Murray, the "father of modern oceanography", who proposed the term radiolarian ooze for the silica deposits of radiolarian shells brought to the surface during the Challenger Expedition.[422] A biogenic ooze is a pelagic sediment containing at least 30 percent from the skeletal remains of marine organisms.
Main types of biogenic ooze
| |||||||
|---|---|---|---|---|---|---|---|
| type | mineral forms |
protist responsible |
name of skeleton |
description | |||
| Siliceous ooze | SiO2 quartz glass opal chert |
diatoms | 
|
frustule | Individual diatoms range in size from 0.002 to 0.2 mm.[423] | ||
| radiolarians | .jpg)
|
skeleton | Radiolarians are protozoa with diameters typically between 0.1 and 0.2 mm that produce intricate mineral skeletons, usually made of silica | ||||
| Calcareous ooze | CaCO3 calcite aragonite limestone chalk |
foraminiferans |
|
test | There are about 10,000 living species of foraminiferans,[424] usually under 1 mm in size. | ||
| coccolithophores | 
|
coccolith | Coccolithophores are spherical cells usually less than 0.1 mm across, enclosed by calcareous plates called coccoliths.[425] Coccoliths are important microfossils. They are the largest global source of biogenic calcium carbonate, and make significant contributions to the global carbon cycle.[426] They are the main constituent of chalk deposits such as the white cliffs of Dover. | ||||
-
 An elaborate mineral skeleton of a radiolarian made of silica.
An elaborate mineral skeleton of a radiolarian made of silica. -
 Diatoms, major components of marine plankton, also have silica skeletons called frustules.
Diatoms, major components of marine plankton, also have silica skeletons called frustules. -
 Coccolithophores have plates or scales made with calcium carbonate called coccoliths
Coccolithophores have plates or scales made with calcium carbonate called coccoliths -

,_Haeckel_(28187768550).jpg)



-
 A diatom microfossil from 40 million years ago
A diatom microfossil from 40 million years ago -
 Diatomaceous earth is a soft, siliceous, sedimentary rock made up of microfossils in the form of the frustules (shells) of single cell diatoms (click to magnify).
Diatomaceous earth is a soft, siliceous, sedimentary rock made up of microfossils in the form of the frustules (shells) of single cell diatoms (click to magnify). -
 Illustration of a Globigerina ooze
Illustration of a Globigerina ooze -
 Shells (tests), usually made of calcium carbonate, from a foraminiferal ooze on the deep ocean floor
Shells (tests), usually made of calcium carbonate, from a foraminiferal ooze on the deep ocean floor




Land interactions
edit
Land interactions impact marine life in many ways. Coastlines typically have continental shelves extending some way from the shore. These provide extensive shallows sunlit down to the seafloor, allowing for photosynthesis and enabling habitats for seagrass meadows, coral reefs, kelp forests and other benthic life. Further from shore the continental shelf slopes towards deep water. Wind blowing at the ocean surface or deep ocean currents can result in cold and nutrient rich waters from abyssal depths moving up the continental slopes. This can result in upwellings along the outer edges of continental shelves, providing conditions for phytoplankton blooms.
Water evaporated by the sun from the surface of the ocean can precipitate on land and eventually return to the ocean as runoff or discharge from rivers, enriched with nutrients as well as pollutants. As rivers discharge into estuaries, freshwater mixes with saltwater and becomes brackish. This provides another shallow water habitat where mangrove forests and estuarine fish thrive. Overall, life in inland lakes can evolve with greater diversity than happens in the sea, because freshwater habitats are themselves diverse and compartmentalised in a way marine habitats are not. Some aquatic life, such as salmon and eels, migrate back and forth between freshwater and marine habitats. These migrations can result in exchanges of pathogens and have impacts on the way life evolves in the ocean.
Anthropogenic impacts
edit
Human activities affect marine life and marine habitats through overfishing, pollution, acidification and the introduction of invasive species. These impact marine ecosystems and food webs and may result in consequences as yet unrecognised for the biodiversity and continuation of marine life forms.[428]
Biodiversity and extinction events
edit

Biodiversity is the result of over three billion years of evolution. Until approximately 600 million years ago, all life consisted of archaea, bacteria, protozoans and similar single-celled organisms. The history of biodiversity during the Phanerozoic (the last 540 million years), starts with rapid growth during the Cambrian explosion – a period during which nearly every phylum of multicellular organisms first appeared. Over the next 400 million years or so, invertebrate diversity showed little overall trend and vertebrate diversity shows an overall exponential trend.[430]
However, more than 99 percent of all species that ever lived on Earth, amounting to over five billion species,[431] are estimated to be extinct.[432][433] These extinctions occur at an uneven rate. The dramatic rise in diversity has been marked by periodic, massive losses of diversity classified as mass extinction events.[430] Mass extinction events occur when life undergoes precipitous global declines. Most diversity and biomass on earth is found among the microorganisms, which are difficult to measure. Recorded extinction events are therefore based on the more easily observed changes in the diversity and abundance of larger multicellular organisms, rather than the total diversity and abundance of life.[434] Marine fossils are mostly used to measure extinction rates because of their superior fossil record and stratigraphic range compared to land organisms.
Based on the fossil record, the background rate of extinctions on Earth is about two to five taxonomic families of marine animals every million years. The Great Oxygenation Event was perhaps the first major extinction event. Since the Cambrian explosion five major mass extinctions have significantly exceeded the background extinction rate.[435] The worst was the Permian-Triassic extinction event, 251 million years ago. One generally estimates that the Big Five mass extinctions of the Phanerozoic (the last 540 million years) wiped out more than 40% of marine genera and probably more than 70% of marine species.[436] The current Holocene extinction caused by human activity, and now referred to as the "sixth extinction", may prove ultimately more devastating.
Investigating and Exploring Marine Life
editResearch and study
editIn order to perform research and enrich Marine Life knowledge, Scientists use various methods in-order to reach and explore the depths of the ocean. several Hi-tech instruments and vehicles are used for this purpose. [437]
- Autonomous Underwater Vehicles (AUVs)- Underwater robots used to explore the ocean. AUVs are independent robots and can explore unmanned. They are released from a ship and are operated from the surface. [438]
- Deep-Towed Vehicles (DTVs)- vehicles towed behind research vessels, offering a simpler alternative to more advanced underwater vehicles. They serve as versatile platforms for deploying oceanographic instruments to measure various ocean parameters, with specific models like the DTV BRIDGET used for studying hydrothermal vent plumes by moving near the ocean floor.[439]
- Manned Submersibles- an manned underwater vehicle used for exploring, experimenting and is often used by army. [437]
- Research vessels (R/Vs)- a boat or ship used to conduct research over a ling period of time. It is capable of transporting a diverse range of sampling and surveying equipment. Research vessels typically feature on-board laboratory space, allowing researchers to promptly analyze the materials collected during cruises.
- Remotely Operated Vehicles (ROVs)- unmanned vehicles. able to reach greater depths under water in order to collect a wider variety of information. [437][440]
See also
edit- Blue Planet – 2001 British nature documentary television series - David Attenborough
- Blue Planet II – 2017 British nature documentary television series
- Census of Marine Life – 10 year international marine biological program
- Colonization of land
- Marine larval ecology
- Taxonomy of invertebrates – System of classification of animals with emphasis on the invertebrates
Notes
edit- ^ This is the measurement taken by the vessel Kaikō in March 1995 and is considered the most accurate measurement to date. See the Challenger Deep article for more details.
- ^ Myxozoa were thought to be an exception, but are now thought to be heavily modified members of the Cnidaria. Jiménez-Guri E, Philippe H, Okamura B, Holland PW (July 2007). "Buddenbrockia is a cnidarian worm". Science. 317 (5834): 116–8. Bibcode:2007Sci...317..116J. doi:10.1126/science.1142024. PMID 17615357. S2CID 5170702.
References
edit- ^ a b Cavicchioli R, Ripple WJ, Timmis KN, Azam F, Bakken LR, Baylis M, et al. (September 2019). "Scientists' warning to humanity: microorganisms and climate change". Nature Reviews. Microbiology. 17 (9): 569–586. doi:10.1038/s41579-019-0222-5. PMC 7136171. PMID 31213707.
 Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ^ a b c "National Oceanic and Atmospheric Administration – Ocean". NOAA. Retrieved 20 February 2019.
- ^ "Tiny Fish May Be Ancestor of Nearly All Living Vertebrates". Live Science. 11 June 2014.
- ^ Drogin, B (2 August 2009). "Mapping an ocean of species". Los Angeles Times. Retrieved 18 August 2009.
- ^ Bouchet, Philippe; Decock, Wim; Lonneville, Britt; Vanhoorne, Bart; Vandepitte, Leen (June 2023). "Marine biodiversity discovery: the metrics of new species descriptions". Frontiers in Marine Science. 10 (3389). doi:10.3389/fmars.2023.929989 – via ResearchGate.
- ^ Paul, GS (2010). "The Evolution of Dinosaurs and their World". The Princeton Field Guide to Dinosaurs. Princeton: Princeton University Press. p. 19. ISBN 978-0-691-13720-9.
- ^ Bortolotti, Dan (2008). Wild blue: a natural history of the world's largest animal. New York: Thomas Dunn Books. ISBN 978-0-312-38387-9. OCLC 213451450.
- ^ a b c Bar-On YM, Phillips R, Milo R (June 2018). "The biomass distribution on Earth". Proceedings of the National Academy of Sciences of the United States of America. 115 (25): 6506–6511. Bibcode:2018PNAS..115.6506B. doi:10.1073/pnas.1711842115. PMC 6016768. PMID 29784790.
- ^ "Census Of Marine Life". Smithsonian. 30 April 2018. Retrieved 29 October 2020.
- ^ Xiao-Feng P (2014). "Chapter 5". Water: Molecular Structure And Properties. World Scientific. pp. 390–461. ISBN 9789814440448.
- ^ Greenwood NN, Earnshaw A (1997). Chemistry of the Elements (2nd ed.). Butterworth-Heinemann. p. 620. ISBN 978-0-08-037941-8.
- ^ "Water, the Universal Solvent". USGS. Archived from the original on 9 July 2017. Retrieved 27 June 2017.
- ^ Reece JB (31 October 2013). Campbell Biology (10 ed.). Pearson. p. 48. ISBN 9780321775658.
- ^ Reece JB (31 October 2013). Campbell Biology (10 ed.). Pearson. p. 44. ISBN 9780321775658.
- ^ Collins JC (1991). The Matrix of Life: A View of Natural Molecules from the Perspective of Environmental Water. Molecular Presentations. ISBN 978-0-9629719-0-7.
- ^ "7,000 m Class Remotely Operated Vehicle KAIKO 7000". Japan Agency for Marine-Earth Science and Technology (JAMSTEC). Archived from the original on 10 April 2020. Retrieved 7 June 2008.
- ^ "How many oceans are there?". NOAA. 9 April 2020.
- ^ Charette MA, Smith WH (June 2010). "The Volume of Earth's Ocean". Oceanography. 23 (2): 112–14. doi:10.5670/oceanog.2010.51. hdl:1912/3862.
- ^ Duxbury AC, Cenedese C (7 May 2021). "Sphere depth of the ocean – hydrology". Encyclopædia Britannica. Retrieved 12 April 2015.
- ^ "Third rock from the Sun – restless Earth". NASA's Cosmos. Retrieved 12 April 2015.
- ^ Perlman H (17 March 2014). "The World's Water". USGS Water-Science School. Retrieved 12 April 2015.
- ^ Kennish KJ (2001). Practical handbook of marine science. Marine science series (3rd ed.). CRC Press. p. 35. ISBN 978-0-8493-2391-1.
- ^ "Why is the ocean salty?".
- ^ Mullen L (11 June 2002). "Salt of the Early Earth". NASA Astrobiology Magazine. Archived from the original on 30 June 2007. Retrieved 14 March 2007.
- ^ Morris RM. "Oceanic Processes". NASA Astrobiology Magazine. Archived from the original on 15 April 2009. Retrieved 14 March 2007.
- ^ Scott M (24 April 2006). "Earth's Big heat Bucket". NASA Earth Observatory. Retrieved 14 March 2007.
- ^ Sample S (21 June 2005). "Sea Surface Temperature". NASA. Archived from the original on 6 April 2013. Retrieved 21 April 2007.
- ^ "Volumes of the World's Oceans from ETOPO1". NOAA. Archived from the original on 11 March 2015. Retrieved 20 February 2019.
{{cite web}}: CS1 maint: bot: original URL status unknown (link) - ^ "Planet "Earth": We Should Have Called It "Sea"". Quote Invertigator. 25 January 2017.
- ^ "Unveiling Planet Ocean". pNASA Science. 14 March 2002. Archived from the original on 8 October 2022. Retrieved 19 February 2019.
- ^ Dyches P, Brown D (12 May 2015). "NASA Research Reveals Europa's Mystery Dark Material Could Be Sea Salt". NASA. Retrieved 12 May 2015.
- ^ Adamu Z (1 October 2012). "Water near surface of a Jupiter moon only temporary". CNN Light Years. Archived from the original on 5 October 2012. Retrieved 24 April 2019.
- ^ Tritt CS (2002). "Possibility of Life on Europa". Milwaukee School of Engineering. Archived from the original on 9 June 2007. Retrieved 10 August 2007.
- ^ Schulze-Makuch D, Irwin LN (2001). "Alternative Energy Sources Could Support Life on Europa" (PDF). Departments of Geological and Biological Sciences, University of Texas at El Paso. Archived from the original (PDF) on 3 July 2006. Retrieved 21 December 2007.
- ^ Friedman L (14 December 2005). "Projects: Europa Mission Campaign". The Planetary Society. Archived from the original on 11 August 2011. Retrieved 8 August 2011.
- ^ "Ocean Within Enceladus May Harbor Hydrothermal Activity". NASA Press Release. 11 March 2015.
- ^ "Age of the Earth". United States Geological Survey. 9 July 2007. Retrieved 31 May 2015.
- ^ Dalrymple GB (January 2001). "The age of the Earth in the twentieth century: a problem (mostly) solved". Special Publications. 190 (1). London: Geological Society: 205–21. Bibcode:2001GSLSP.190..205D. doi:10.1144/GSL.SP.2001.190.01.14. S2CID 130092094.
- ^ Manhes G, Allègre CJ, Dupré B, Hamelin B (May 1980). "Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics". Earth and Planetary Science Letters. 47 (3): 370–382. Bibcode:1980E&PSL..47..370M. doi:10.1016/0012-821X(80)90024-2. ISSN 0012-821X.
- ^ Schopf JW, Kudryavtsev AB, Czaja AD, Tripathi AB (5 October 2007). "Evidence of Archean life: Stromatolites and microfossils". Precambrian Research. 158 (3–4): 141–155. Bibcode:2007PreR..158..141S. doi:10.1016/j.precamres.2007.04.009. ISSN 0301-9268.
- ^ Raven PH, Johnson GB (2002). Biology (6th ed.). Boston: McGraw-Hill. p. 68. ISBN 978-0-07-112261-0.
- ^ Baumgartner RJ, Van Kranendonk MJ, Wacey D, Fiorentini ML, Saunders M, Caruso S, et al. (2019). "Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life" (PDF). Geology. 47 (11): 1039–1043. Bibcode:2019Geo....47.1039B. doi:10.1130/G46365.1. S2CID 204258554.
- ^ "Earliest signs of life: Scientists find microbial remains in ancient rocks". Phys.org. 26 September 2019.
- ^ Ohtomo Y, Kakegawa T, Ishida A, Nagase T, Rosing MT (January 2014). "Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks". Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe...7...25O. doi:10.1038/ngeo2025. ISSN 1752-0894.
- ^ a b Borenstein S (19 October 2015). "Hints of life on what was thought to be desolate early Earth". Associated Press. Retrieved 9 October 2018.
- ^ Bell EA, Boehnke P, Harrison TM, Mao WL (November 2015). "Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon". Proceedings of the National Academy of Sciences of the United States of America. 112 (47): 14518–21. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481.
- ^ Penny D, Poole A (December 1999). "The nature of the last universal common ancestor". Current Opinion in Genetics & Development. 9 (6): 672–7. doi:10.1016/S0959-437X(99)00020-9. PMID 10607605.
- ^ Theobald DL (May 2010). "A formal test of the theory of universal common ancestry". Nature. 465 (7295): 219–22. Bibcode:2010Natur.465..219T. doi:10.1038/nature09014. PMID 20463738. S2CID 4422345.
- ^ Doolittle WF (February 2000). "Uprooting the tree of life" (PDF). Scientific American. 282 (2): 90–5. Bibcode:2000SciAm.282b..90D. doi:10.1038/scientificamerican0200-90. PMID 10710791. Archived from the original (PDF) on 7 September 2006. Retrieved 5 April 2015.
- ^ Peretó J (March 2005). "Controversies on the origin of life" (PDF). International Microbiology. 8 (1): 23–31. PMID 15906258. Archived from the original (PDF) on 24 August 2015.
- ^ Joyce GF (July 2002). "The antiquity of RNA-based evolution". Nature. 418 (6894): 214–21. Bibcode:2002Natur.418..214J. doi:10.1038/418214a. PMID 12110897. S2CID 4331004.
- ^ Trevors JT, Psenner R (December 2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". FEMS Microbiology Reviews. 25 (5): 573–82. doi:10.1111/j.1574-6976.2001.tb00592.x. PMID 11742692.
- ^ Wade N (25 July 2016). "Meet Luca, the Ancestor of All Living Things". New York Times. Retrieved 25 July 2016.
- ^ Bapteste E, Walsh DA (June 2005). "Does the 'Ring of Life' ring true?". Trends in Microbiology. 13 (6): 256–61. doi:10.1016/j.tim.2005.03.012. PMID 15936656.
- ^ Darwin C (1859). "On The Origin of the Species". London: John Murray.
- ^ Doolittle WF, Bapteste E (February 2007). "Pattern pluralism and the Tree of Life hypothesis". Proceedings of the National Academy of Sciences of the United States of America. 104 (7): 2043–9. Bibcode:2007PNAS..104.2043D. doi:10.1073/pnas.0610699104. PMC 1892968. PMID 17261804.
- ^ Kunin V, Goldovsky L, Darzentas N, Ouzounis CA (July 2005). "The net of life: reconstructing the microbial phylogenetic network". Genome Research. 15 (7): 954–9. doi:10.1101/gr.3666505. PMC 1172039. PMID 15965028.
- ^ Jablonski D (June 1999). "The future of the fossil record". Science. 284 (5423): 2114–6. doi:10.1126/science.284.5423.2114. PMID 10381868. S2CID 43388925.
- ^ Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P (March 2006). "Toward automatic reconstruction of a highly resolved tree of life". Science. 311 (5765): 1283–7. Bibcode:2006Sci...311.1283C. CiteSeerX 10.1.1.381.9514. doi:10.1126/science.1123061. PMID 16513982. S2CID 1615592.
- ^ Mason SF (6 September 1984). "Origins of biomolecular handedness". Nature. 311 (5981): 19–23. Bibcode:1984Natur.311...19M. doi:10.1038/311019a0. PMID 6472461. S2CID 103653.
- ^ Wolf YI, Rogozin IB, Grishin NV, Koonin EV (September 2002). "Genome trees and the tree of life". Trends in Genetics. 18 (9): 472–9. doi:10.1016/S0168-9525(02)02744-0. PMID 12175808.
- ^ Varki A, Altheide TK (December 2005). "Comparing the human and chimpanzee genomes: searching for needles in a haystack". Genome Research. 15 (12): 1746–58. doi:10.1101/gr.3737405. PMID 16339373.
- ^ a b Cavalier-Smith T (June 2006). "Cell evolution and Earth history: stasis and revolution". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. PMC 1578732. PMID 16754610.
- ^ Schopf JW (June 2006). "Fossil evidence of Archaean life". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 361 (1470): 869–85. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
- Altermann W, Kazmierczak J (November 2003). "Archean microfossils: a reappraisal of early life on Earth". Research in Microbiology. 154 (9): 611–7. doi:10.1016/j.resmic.2003.08.006. PMID 14596897.
- ^ a b Schopf JW (July 1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proceedings of the National Academy of Sciences of the United States of America. 91 (15): 6735–42. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691.
- ^ a b Poole AM, Penny D (January 2007). "Evaluating hypotheses for the origin of eukaryotes". BioEssays. 29 (1): 74–84. doi:10.1002/bies.20516. PMID 17187354.
- ^ a b Dyall SD, Brown MT, Johnson PJ (April 2004). "Ancient invasions: from endosymbionts to organelles". Science. 304 (5668): 253–7. Bibcode:2004Sci...304..253D. doi:10.1126/science.1094884. PMID 15073369. S2CID 19424594.
- ^ Martin W (October 2005). "The missing link between hydrogenosomes and mitochondria". Trends in Microbiology. 13 (10): 457–9. doi:10.1016/j.tim.2005.08.005. PMID 16109488.
- ^ Lang BF, Gray MW, Burger G (December 1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annual Review of Genetics. 33: 351–97. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- McFadden GI (December 1999). "Endosymbiosis and evolution of the plant cell". Current Opinion in Plant Biology. 2 (6): 513–9. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ DeLong EF, Pace NR (August 2001). "Environmental diversity of bacteria and archaea". Systematic Biology. 50 (4): 470–8. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Kaiser D (December 2001). "Building a multicellular organism". Annual Review of Genetics. 35: 103–23. doi:10.1146/annurev.genet.35.102401.090145. PMID 11700279. S2CID 18276422.
- ^ Zimmer C (7 January 2016). "Genetic Flip Helped Organisms Go From One Cell to Many". The New York Times. Retrieved 7 January 2016.
- ^ Valentine JW, Jablonski D, Erwin DH (February 1999). "Fossils, molecules and embryos: new perspectives on the Cambrian explosion". Development. 126 (5): 851–9. doi:10.1242/dev.126.5.851. PMID 9927587.
- ^ Ohno S (January 1997). "The reason for as well as the consequence of the Cambrian explosion in animal evolution". Journal of Molecular Evolution. 44 (Suppl. 1): S23-7. Bibcode:1997JMolE..44S..23O. doi:10.1007/PL00000055. PMID 9071008. S2CID 21879320.
- Valentine JW, Jablonski D (2003). "Morphological and developmental macroevolution: a paleontological perspective". The International Journal of Developmental Biology. 47 (7–8): 517–22. PMID 14756327. Retrieved 30 December 2014.
- ^ Wellman CH, Osterloff PL, Mohiuddin U (September 2003). "Fragments of the earliest land plants" (PDF). Nature. 425 (6955): 282–5. Bibcode:2003Natur.425..282W. doi:10.1038/nature01884. PMID 13679913. S2CID 4383813.
- ^ Barton N (2007). Evolution. CSHL Press. pp. 273–274. ISBN 9780199226320. Retrieved 30 September 2012.
- ^ Waters ER (December 2003). "Molecular adaptation and the origin of land plants". Molecular Phylogenetics and Evolution. 29 (3): 456–63. doi:10.1016/j.ympev.2003.07.018. PMID 14615186.
- ^ Mayhew PJ (August 2007). "Why are there so many insect species? Perspectives from fossils and phylogenies". Biological Reviews of the Cambridge Philosophical Society. 82 (3): 425–54. doi:10.1111/j.1469-185X.2007.00018.x. PMID 17624962. S2CID 9356614.
- ^ Carroll RL (May 2007). "The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians". Zoological Journal of the Linnean Society. 150 (Supplement s1): 1–140. doi:10.1111/j.1096-3642.2007.00246.x. ISSN 1096-3642.
- ^ Wible JR, Rougier GW, Novacek MJ, Asher RJ (June 2007). "Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary". Nature. 447 (7147): 1003–6. Bibcode:2007Natur.447.1003W. doi:10.1038/nature05854. PMID 17581585. S2CID 4334424.
- ^ Witmer LM (July 2011). "Palaeontology: An icon knocked from its perch". Nature. 475 (7357): 458–9. doi:10.1038/475458a. PMID 21796198. S2CID 205066360.
- ^ Schloss PD, Handelsman J (December 2004). "Status of the microbial census". Microbiology and Molecular Biology Reviews. 68 (4): 686–91. doi:10.1128/MMBR.68.4.686-691.2004. PMC 539005. PMID 15590780.
- ^ Miller GT, Spoolman S (January 2012). "Chapter 4.1: What is Biodiversity and Why is it Important?". Environmental Science. Cengage Learning. p. 62. ISBN 978-1-133-70787-5.
- ^ Mora C, Tittensor DP, Adl S, Simpson AG, Worm B (August 2011). "How many species are there on Earth and in the ocean?". PLOS Biology. 9 (8): e1001127. doi:10.1371/journal.pbio.1001127. PMC 3160336. PMID 21886479.
- ^ Madigan M, Martinko J, eds. (2006). Brock Biology of Microorganisms (13th ed.). Pearson Education. p. 1096. ISBN 978-0-321-73551-5.
- ^ Rybicki EP (1990). "The classification of organisms at the edge of life, or problems with virus systematics". South African Journal of Science. 86: 182–6. ISSN 0038-2353.
- ^ Lwoff A (October 1957). "The concept of virus". Journal of General Microbiology. 17 (2): 239–53. doi:10.1099/00221287-17-2-239. PMID 13481308.
- ^ "2002 WHO mortality data". Archived from the original on 19 August 2006. Retrieved 20 January 2007.
- ^ "Functions of global ocean microbiome key to understanding environmental changes". www.sciencedaily.com. University of Georgia. 10 December 2015. Retrieved 11 December 2015.
- ^ Suttle CA (September 2005). "Viruses in the sea". Nature. 437 (7057): 356–61. Bibcode:2005Natur.437..356S. doi:10.1038/nature04160. PMID 16163346. S2CID 4370363.
- ^ a b Shors T (2017). Understanding Viruses (3rd ed.). Jones and Bartlett Publishers. ISBN 978-1-284-02592-7.
- ^ a b c Suttle CA (October 2007). "Marine viruses--major players in the global ecosystem". Nature Reviews. Microbiology. 5 (10): 801–12. doi:10.1038/nrmicro1750. PMID 17853907. S2CID 4658457.
- ^ Morrison J (11 January 2016). "Living Bacteria Are Riding Earth's Air Currents". Smithsonian Magazine.
- ^ Robbins J (13 April 2018). "Trillions Upon Trillions of Viruses Fall From the Sky Each Day". The New York Times. Retrieved 14 April 2018.
- ^ Reche I, D'Orta G, Mladenov N, Winget DM, Suttle CA (April 2018). "Deposition rates of viruses and bacteria above the atmospheric boundary layer". The ISME Journal. 12 (4): 1154–1162. doi:10.1038/s41396-017-0042-4. PMC 5864199. PMID 29379178.
- ^ Staff (2014). "The Biosphere". Aspen Global Change Institute. Archived from the original on 2 September 2010. Retrieved 10 November 2014.
- ^ a b c Choi CQ (17 March 2013). "Microbes Thrive in Deepest Spot on Earth". LiveScience. Retrieved 17 March 2013.
- ^ Glud RN, Wenzhöfer F, Middelboe M, Oguri K, Turnewitsch R, Canfield DE, Kitazato H (17 March 2013). "High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth". Nature Geoscience. 6 (4): 284–288. Bibcode:2013NatGe...6..284G. doi:10.1038/ngeo1773.
- ^ Oskin B (14 March 2013). "Intraterrestrials: Life Thrives in Ocean Floor". LiveScience. Retrieved 17 March 2013.
- ^ Morelle R (15 December 2014). "Microbes discovered by deepest marine drill analysed". BBC News. Retrieved 15 December 2014.
- ^ Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, Miyazaki J, et al. (August 2008). "Cell proliferation at 122 degrees C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation". Proceedings of the National Academy of Sciences of the United States of America. 105 (31): 10949–54. Bibcode:2008PNAS..10510949T. doi:10.1073/pnas.0712334105. PMC 2490668. PMID 18664583.
- ^ Fox D (August 2014). "Lakes under the ice: Antarctica's secret garden". Nature. 512 (7514): 244–6. Bibcode:2014Natur.512..244F. doi:10.1038/512244a. PMID 25143097.
- ^ Mack E (20 August 2014). "Life Confirmed Under Antarctic Ice; Is Space Next?". Forbes. Retrieved 21 August 2014.
- ^ Wimmer E, Mueller S, Tumpey TM, Taubenberger JK (December 2009). "Synthetic viruses: a new opportunity to understand and prevent viral disease". Nature Biotechnology. 27 (12): 1163–72. doi:10.1038/nbt.1593. PMC 2819212. PMID 20010599.
- ^ Koonin EV, Senkevich TG, Dolja VV (September 2006). "The ancient Virus World and evolution of cells". Biology Direct. 1: 29. doi:10.1186/1745-6150-1-29. PMC 1594570. PMID 16984643.
- ^ Topley WW, Wilson GS, Collier LH, Balows A, Sussman M (1998). Mahy BW, Collier L (eds.). Topley and Wilson's Microbiology and Microbial Infections. Vol. 1 (9th ed.). London: Arnold. pp. 33–37. ISBN 978-0-340-66316-5.
- ^ Iyer LM, Balaji S, Koonin EV, Aravind L (April 2006). "Evolutionary genomics of nucleo-cytoplasmic large DNA viruses". Virus Research. 117 (1): 156–84. doi:10.1016/j.virusres.2006.01.009. PMID 16494962.
- ^ Sanjuán R, Nebot MR, Chirico N, Mansky LM, Belshaw R (October 2010). "Viral mutation rates". Journal of Virology. 84 (19): 9733–48. doi:10.1128/JVI.00694-10. PMC 2937809. PMID 20660197.
- ^ Mahy WJ, Van Regenmortel MH, eds. (2009). Desk Encyclopedia of General Virology. Oxford: Academic Press. p. 28. ISBN 978-0-12-375146-1.
- ^ a b Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H (August 2003). "Phage as agents of lateral gene transfer". Current Opinion in Microbiology. 6 (4): 417–24. doi:10.1016/S1369-5274(03)00086-9. PMID 12941415.
- ^ a b Koonin EV, Starokadomskyy P (October 2016). "Are viruses alive? The replicator paradigm sheds decisive light on an old but misguided question". Studies in History and Philosophy of Biological and Biomedical Sciences. 59: 125–34. doi:10.1016/j.shpsc.2016.02.016. PMC 5406846. PMID 26965225.
- ^ Rybicki EP (1990). "The classification of organisms at the edge of life, or problems with virus systematics". South African Journal of Science. 86: 182–186.
- ^ a b Mann NH (May 2005). "The third age of phage". PLOS Biology. 3 (5): e182. doi:10.1371/journal.pbio.0030182. PMC 1110918. PMID 15884981.
- ^ Wommack KE, Colwell RR (March 2000). "Virioplankton: viruses in aquatic ecosystems". Microbiology and Molecular Biology Reviews. 64 (1): 69–114. doi:10.1128/MMBR.64.1.69-114.2000. PMC 98987. PMID 10704475.
- ^ a b Suttle CA (September 2005). "Viruses in the sea". Nature. 437 (7057): 356–61. Bibcode:2005Natur.437..356S. doi:10.1038/nature04160. PMID 16163346. S2CID 4370363.
- ^ Bergh O, Børsheim KY, Bratbak G, Heldal M (August 1989). "High abundance of viruses found in aquatic environments". Nature. 340 (6233): 467–8. Bibcode:1989Natur.340..467B. doi:10.1038/340467a0. PMID 2755508. S2CID 4271861.
- ^ Wigington CH, Sonderegger D, Brussaard CP, Buchan A, Finke JF, Fuhrman JA, et al. (January 2016). "Re-examination of the relationship between marine virus and microbial cell abundances" (PDF). Nature Microbiology. 1 (3): 15024. doi:10.1038/nmicrobiol.2015.24. PMID 27572161. S2CID 52829633.
- ^ Krupovic M, Bamford DH (July 2007). "Putative prophages related to lytic tailless marine dsDNA phage PM2 are widespread in the genomes of aquatic bacteria". BMC Genomics. 8: 236. doi:10.1186/1471-2164-8-236. PMC 1950889. PMID 17634101.
- ^ Xue H, Xu Y, Boucher Y, Polz MF (January 2012). "High frequency of a novel filamentous phage, VCY φ, within an environmental Vibrio cholerae population". Applied and Environmental Microbiology. 78 (1): 28–33. Bibcode:2012ApEnM..78...28X. doi:10.1128/AEM.06297-11. PMC 3255608. PMID 22020507.
- ^ Roux S, Krupovic M, Poulet A, Debroas D, Enault F (2012). "Evolution and diversity of the Microviridae viral family through a collection of 81 new complete genomes assembled from virome reads". PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO...740418R. doi:10.1371/journal.pone.0040418. PMC 3394797. PMID 22808158.
- ^ "Harmful Algal Blooms: Red Tide: Home |CDC HSB". www.cdc.gov. Retrieved 19 December 2014.
- ^ Lawrence CM, Menon S, Eilers BJ, Bothner B, Khayat R, Douglas T, Young MJ (May 2009). "Structural and functional studies of archaeal viruses". The Journal of Biological Chemistry. 284 (19): 12599–603. doi:10.1074/jbc.R800078200. PMC 2675988. PMID 19158076.
- ^ Prangishvili D, Forterre P, Garrett RA (November 2006). "Viruses of the Archaea: a unifying view". Nature Reviews. Microbiology. 4 (11): 837–48. doi:10.1038/nrmicro1527. PMID 17041631. S2CID 9915859.
- ^ Prangishvili D, Garrett RA (April 2004). "Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses" (PDF). Biochemical Society Transactions. 32 (Pt 2): 204–8. doi:10.1042/BST0320204. PMID 15046572. S2CID 20018642.
- ^ Forterre P, Philippe H (June 1999). "The last universal common ancestor (LUCA), simple or complex?". The Biological Bulletin. 196 (3): 373–5, discussion 375–7. doi:10.2307/1542973. JSTOR 1542973. PMID 11536914.
- ^ Fredrickson JK, Zachara JM, Balkwill DL, Kennedy D, Li SM, Kostandarithes HM, et al. (July 2004). "Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the hanford site, washington state". Applied and Environmental Microbiology. 70 (7): 4230–41. Bibcode:2004ApEnM..70.4230F. doi:10.1128/AEM.70.7.4230-4241.2004. PMC 444790. PMID 15240306.
- ^ Woese CR, Kandler O, Wheelis ML (June 1990). "Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya". Proceedings of the National Academy of Sciences of the United States of America. 87 (12): 4576–9. Bibcode:1990PNAS...87.4576W. doi:10.1073/pnas.87.12.4576. PMC 54159. PMID 2112744.
- ^ DeLong EF, Pace NR (August 2001). "Environmental diversity of bacteria and archaea". Systematic Biology. 50 (4): 470–8. CiteSeerX 10.1.1.321.8828. doi:10.1080/106351501750435040. PMID 12116647.
- ^ Brown JR, Doolittle WF (December 1997). "Archaea and the prokaryote-to-eukaryote transition". Microbiology and Molecular Biology Reviews. 61 (4): 456–502. doi:10.1128/mmbr.61.4.456-502.1997. PMC 232621. PMID 9409149.
- ^ Lang BF, Gray MW, Burger G (1999). "Mitochondrial genome evolution and the origin of eukaryotes". Annual Review of Genetics. 33: 351–97. doi:10.1146/annurev.genet.33.1.351. PMID 10690412.
- ^ McFadden GI (December 1999). "Endosymbiosis and evolution of the plant cell". Current Opinion in Plant Biology. 2 (6): 513–9. doi:10.1016/S1369-5266(99)00025-4. PMID 10607659.
- ^ Keeling PJ (2004). "Diversity and evolutionary history of plastids and their hosts". American Journal of Botany. 91 (10): 1481–1493. doi:10.3732/ajb.91.10.1481. PMID 21652304. S2CID 17522125.
- ^ "The largest Bacterium: Scientist discovers new bacterial life form off the African coast", Max Planck Institute for Marine Microbiology, 8 April 1999, archived from the original on 20 January 2010
- ^ List of Prokaryotic names with Standing in Nomenclature - Genus Thiomargarita
- ^ Bang C, Schmitz RA (September 2015). "Archaea associated with human surfaces: not to be underestimated". FEMS Microbiology Reviews. 39 (5): 631–48. doi:10.1093/femsre/fuv010. PMID 25907112.
- ^ "Archaea". Online Etymology Dictionary. Retrieved 17 August 2016.
- ^ Pace NR (May 2006). "Time for a change". Nature. 441 (7091): 289. Bibcode:2006Natur.441..289P. doi:10.1038/441289a. PMID 16710401. S2CID 4431143.
- ^ Stoeckenius W (October 1981). "Walsby's square bacterium: fine structure of an orthogonal procaryote". Journal of Bacteriology. 148 (1): 352–60. doi:10.1128/JB.148.1.352-360.1981. PMC 216199. PMID 7287626.
- ^ Whittaker RH, Margulis L (April 1978). "Protist classification and the kingdoms of organisms". Bio Systems. 10 (1–2): 3–18. doi:10.1016/0303-2647(78)90023-0. PMID 418827.
- ^ Faure E, Not F, Benoiston AS, Labadie K, Bittner L, Ayata SD (April 2019). "Mixotrophic protists display contrasted biogeographies in the global ocean". The ISME Journal. 13 (4): 1072–1083. doi:10.1038/s41396-018-0340-5. PMC 6461780. PMID 30643201.
- ^ a b Leles SG, Mitra A, Flynn KJ, Stoecker DK, Hansen PJ, Calbet A, et al. (August 2017). "Oceanic protists with different forms of acquired phototrophy display contrasting biogeographies and abundance". Proceedings. Biological Sciences. 284 (1860): 20170664. doi:10.1098/rspb.2017.0664. PMC 5563798. PMID 28768886.
- ^ Budd GE, Jensen S (February 2017). "The origin of the animals and a 'Savannah' hypothesis for early bilaterian evolution". Biological Reviews of the Cambridge Philosophical Society. 92 (1): 446–473. doi:10.1111/brv.12239. PMID 26588818.
- ^ Cavalier-Smith T (December 1993). "Kingdom protozoa and its 18 phyla". Microbiological Reviews. 57 (4): 953–94. doi:10.1128/MMBR.57.4.953-994.1993. PMC 372943. PMID 8302218.
- ^ Corliss JO (1992). "Should there be a separate code of nomenclature for the protists?". Bio Systems. 28 (1–3): 1–14. doi:10.1016/0303-2647(92)90003-H. PMID 1292654.
- ^ Slapeta J, Moreira D, López-García P (October 2005). "The extent of protist diversity: insights from molecular ecology of freshwater eukaryotes". Proceedings. Biological Sciences. 272 (1576): 2073–81. doi:10.1098/rspb.2005.3195. PMC 1559898. PMID 16191619.
- ^ Moreira D, López-García P (January 2002). "The molecular ecology of microbial eukaryotes unveils a hidden world". Trends in Microbiology. 10 (1): 31–8. doi:10.1016/S0966-842X(01)02257-0. PMID 11755083.
- ^ a b Alverson A (11 June 2014). "The Air You're Breathing? A Diatom Made That". Live Science.
- ^ "More on Diatoms". University of California Museum of Paleontology. Archived from the original on 4 October 2012. Retrieved 27 June 2019.
- ^ Devreotes P (September 1989). "Dictyostelium discoideum: a model system for cell-cell interactions in development". Science. 245 (4922): 1054–8. Bibcode:1989Sci...245.1054D. doi:10.1126/science.2672337. PMID 2672337.
- ^ Matz MV, Frank TM, Marshall NJ, Widder EA, Johnsen S (December 2008). "Giant deep-sea protist produces bilaterian-like traces". Current Biology. 18 (23). Elsevier Ltd: 1849–54. doi:10.1016/j.cub.2008.10.028. PMID 19026540. S2CID 8819675.
- ^ Gooday AJ, Da Silva AA, Pawlowski J (1 December 2011). "Xenophyophores (Rhizaria, Foraminifera) from the Nazaré Canyon (Portuguese margin, NE Atlantic)". Deep-Sea Research Part II: Topical Studies in Oceanography. The Geology, Geochemistry, and Biology of Submarine Canyons West of Portugal. 58 (23–24): 2401–2419. Bibcode:2011DSRII..58.2401G. doi:10.1016/j.dsr2.2011.04.005.
- ^ Neil AC, Reece JB, Simon EJ (2004). Essential biology with physiology. Pearson/Benjamin Cummings. p. 291. ISBN 978-0-8053-7503-9.
- ^ O'Malley MA, Simpson AG, Roger AJ (2012). "The other eukaryotes in light of evolutionary protistology". Biology & Philosophy. 28 (2): 299–330. doi:10.1007/s10539-012-9354-y. S2CID 85406712.
- ^ Adl SM, Simpson AG, Farmer MA, Andersen RA, Anderson OR, Barta JR, et al. (2005). "The new higher level classification of eukaryotes with emphasis on the taxonomy of protists". The Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111/j.1550-7408.2005.00053.x. PMID 16248873. S2CID 8060916.
- ^ Margulis L, Chapman MJ (19 March 2009). Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth. Academic Press. ISBN 9780080920146.
- ^ Fang J (April 2010). "Animals thrive without oxygen at sea bottom". Nature. 464 (7290): 825. Bibcode:2010Natur.464..825F. doi:10.1038/464825b. PMID 20376121.
- ^ "Briny deep basin may be home to animals thriving without oxygen". Science News. 23 September 2013.
- ^ Jónasdóttir, Sigrún Huld; Visser, André W.; Richardson, Katherine; Heath, Michael R. (29 September 2015). "Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic". Proceedings of the National Academy of Sciences. 112 (39): 12122–12126. doi:10.1073/pnas.1512110112. ISSN 0027-8424. PMC 4593097. PMID 26338976.
- ^ Pinti, Jérôme; Jónasdóttir, Sigrún H.; Record, Nicholas R.; Visser, André W. (7 March 2023). "The global contribution of seasonally migrating copepods to the biological carbon pump". Limnology and Oceanography. 68 (5): 1147–1160. Bibcode:2023LimOc..68.1147P. doi:10.1002/lno.12335. ISSN 0024-3590. S2CID 257422956.
- ^ Pepato, Almir R.; Vidigal, Teofânia H.D.A.; Klimov, Pavel B. (2018). "Molecular phylogeny of marine mites (Acariformes: Halacaridae), the oldest radiation of extant secondarily marine animals". Molecular Phylogenetics and Evolution. 129: 182–188. doi:10.1016/j.ympev.2018.08.012. PMID 30172010. S2CID 52145427.
- ^ Hyde KD, Jones EG, Leaño E, Pointing SB, Poonyth AD, Vrijmoed LL (1998). "Role of fungi in marine ecosystems". Biodiversity and Conservation. 7 (9): 1147–1161. doi:10.1023/A:1008823515157. S2CID 22264931.
- ^ Kirk PM, Cannon PF, Minter DW, Stalpers J (2008). Dictionary of the Fungi (10 ed.). CABI.
- ^ Hyde KD, Greenwood R, Jones EG (1989). "Spore attachment in marine fungi". Botanica Marina. 32 (3): 205–218. doi:10.1515/botm.1989.32.3.205. S2CID 84879817.
- ^ Le Calvez T, Burgaud G, Mahé S, Barbier G, Vandenkoornhuyse P (October 2009). "Fungal diversity in deep-sea hydrothermal ecosystems". Applied and Environmental Microbiology. 75 (20): 6415–21. Bibcode:2009ApEnM..75.6415L. doi:10.1128/AEM.00653-09. PMC 2765129. PMID 19633124.
- ^ San-Martin A, Orejarena S, Gallardo C, Silva M, Becerra J, Reinoso RO, Chamy MC, Vergara K, Rovirosa J (2008). "Steroids from the marine fungus Geotrichum sp". Journal of the Chilean Chemical Society. 53 (1): 1377–1378. doi:10.4067/S0717-97072008000100011.
- ^ Jones EB, Hyde KD, Pang KL, eds. (2014). Freshwater fungi: and fungal-like organisms. Berlin/Boston: De Gruyter. ISBN 9783110333480.
- ^ Jones EB, Pang KL, eds. (2012). Marine Fungi, and Fungal-like Organisms. Marine and Freshwater Botany. Berlin, Boston: De Gruyter (published August 2012). doi:10.1515/9783110264067. ISBN 978-3-11-026406-7. Retrieved 3 September 2015.
- ^ Wang X, Singh P, Gao Z, Zhang X, Johnson ZI, Wang G (2014). "Distribution and diversity of planktonic fungi in the West Pacific Warm Pool". PLOS ONE. 9 (7): e101523. Bibcode:2014PLoSO...9j1523W. doi:10.1371/journal.pone.0101523.s001. PMC 4081592. PMID 24992154.
- ^ Wang G, Wang X, Liu X, Li Q (2012). "Diversity and biogeochemical function of planktonic fungi in the ocean". In Raghukumar C (ed.). Biology of marine fungi. Progress in Molecular and Subcellular Biology. Vol. 53. Berlin, Heidelberg: Springer-Verlag. pp. 71–88. doi:10.1007/978-3-642-23342-5. ISBN 978-3-642-23341-8. S2CID 39378040. Retrieved 3 September 2015.
- ^ Damare S, Raghukumar C (July 2008). "Fungi and macroaggregation in deep-sea sediments". Microbial Ecology. 56 (1): 168–77. doi:10.1007/s00248-007-9334-y. PMID 17994287. S2CID 21288251.
- ^ Kubanek J, Jensen PR, Keifer PA, Sullards MC, Collins DO, Fenical W (June 2003). "Seaweed resistance to microbial attack: a targeted chemical defense against marine fungi". Proceedings of the National Academy of Sciences of the United States of America. 100 (12): 6916–21. Bibcode:2003PNAS..100.6916K. doi:10.1073/pnas.1131855100. PMC 165804. PMID 12756301.
- ^ a b Gao Z, Johnson ZI, Wang G (January 2010). "Molecular characterization of the spatial diversity and novel lineages of mycoplankton in Hawaiian coastal waters". The ISME Journal. 4 (1): 111–20. doi:10.1038/ismej.2009.87. PMID 19641535.
- ^ Panzer K, Yilmaz P, Weiß M, Reich L, Richter M, Wiese J, et al. (30 July 2015). "Identification of Habitat-Specific Biomes of Aquatic Fungal Communities Using a Comprehensive Nearly Full-Length 18S rRNA Dataset Enriched with Contextual Data". PLOS ONE. 10 (7): e0134377. Bibcode:2015PLoSO..1034377P. doi:10.1371/journal.pone.0134377. PMC 4520555. PMID 26226014.
- ^ Gutierrez MH, Pantoja S, Quinones RA, Gonzalez RR (2010). "Primer registro de hongos filamentosos en el ecosistema de surgencia costero frente a Chile central" [First record of filamentous fungi in the coastal upwelling ecosystem off central Chile]. Gayana (in Spanish). 74 (1): 66–73.
- ^ a b Sridhar KR (2009). "10. Aquatic fungi – Are they planktonic?". Plankton Dynamics of Indian Waters. Jaipur, India: Pratiksha Publications. pp. 133–148.
- ^ "Species of Higher Marine Fungi". University of Mississippi. Archived from the original on 22 April 2013. Retrieved 5 February 2012.
- ^ Hawksworth DL (2000). "Freshwater and marine lichen-forming fungi" (PDF). Fungal Diversity. 5: 1–7.
- ^ "Lichens". National Park Service, US Department of the Interior, Government of the United States. 22 May 2016. Retrieved 4 April 2018.
- ^ a b "The Earth Life Web, Growth and Development in Lichens". earthlife.net. 14 February 2020.
- ^ Silliman BR, Newell SY (December 2003). "Fungal farming in a snail". Proceedings of the National Academy of Sciences of the United States of America. 100 (26): 15643–8. Bibcode:2003PNAS..10015643S. doi:10.1073/pnas.2535227100. PMC 307621. PMID 14657360.
- ^ Yuan X, Xiao S, Taylor TN (May 2005). "Lichen-like symbiosis 600 million years ago". Science. 308 (5724): 1017–20. Bibcode:2005Sci...308.1017Y. doi:10.1126/science.1111347. PMID 15890881. S2CID 27083645.
- ^ Jones EB, Pang KL (31 August 2012). Marine Fungi: and Fungal-like Organisms. Walter de Gruyter. ISBN 9783110264067.
- ^ Davidson MW (26 May 2005). "Animal Cell Structure". Molecular Expressions. Tallahassee, Fla.: Florida State University. Retrieved 3 September 2008.
- ^ Vogel G (20 September 2018). "This fossil is one of the world's earliest animals, according to fat molecules preserved for a half-billion years". Science. AAAS. Retrieved 21 September 2018.
- ^ Bobrovskiy I, Hope JM, Ivantsov A, Nettersheim BJ, Hallmann C, Brocks JJ (September 2018). "Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals". Science. 361 (6408): 1246–1249. Bibcode:2018Sci...361.1246B. doi:10.1126/science.aat7228. hdl:1885/230014. PMID 30237355.
- ^ Retallack GJ (2007). "Growth, decay and burial compaction of Dickinsonia, an iconic Ediacaran fossil" (PDF). Alcheringa: An Australasian Journal of Palaeontology. 31 (3): 215–240. Bibcode:2007Alch...31..215R. doi:10.1080/03115510701484705. S2CID 17181699.
- ^ Sperling EA, Vinther J (2010). "A placozoan affinity for Dickinsonia and the evolution of late Proterozoic metazoan feeding modes". Evolution & Development. 12 (2): 201–9. doi:10.1111/j.1525-142X.2010.00404.x. PMID 20433459. S2CID 38559058.
- ^ Gold DA, Runnegar B, Gehling JG, Jacobs DK (2015). "Ancestral state reconstruction of ontogeny supports a bilaterian affinity for Dickinsonia". Evolution & Development. 17 (6): 315–24. doi:10.1111/ede.12168. PMID 26492825. S2CID 26099557.
- ^ Chen JY, Oliveri P, Gao F, Dornbos SQ, Li CW, Bottjer DJ, Davidson EH (August 2002). "Precambrian animal life: probable developmental and adult cnidarian forms from Southwest China" (PDF). Developmental Biology. 248 (1): 182–96. doi:10.1006/dbio.2002.0714. PMID 12142030. Archived from the original (PDF) on 26 May 2013. Retrieved 4 February 2015.
- ^ Grazhdankin D (June 2004). "Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution". Paleobiology. 30 (2): 203–221. doi:10.1666/0094-8373(2004)030<0203:PODITE>2.0.CO;2. ISSN 0094-8373. S2CID 129376371.
- ^ Seilacher A (August 1992). "Vendobionta and Psammocorallia: lost constructions of Precambrian evolution". Journal of the Geological Society. 149 (4): 607–613. Bibcode:1992JGSoc.149..607S. doi:10.1144/gsjgs.149.4.0607. ISSN 0016-7649. S2CID 128681462. Retrieved 4 February 2015.
- ^ Martin MW, Grazhdankin DV, Bowring SA, Evans DA, Fedonkin MA, Kirschvink JL (May 2000). "Age of Neoproterozoic bilatarian body and trace fossils, White Sea, Russia: implications for metazoan evolution". Science. 288 (5467): 841–5. Bibcode:2000Sci...288..841M. doi:10.1126/science.288.5467.841. PMID 10797002. S2CID 1019572.
- ^ Fedonkin MA, Waggoner BM (28 August 1997). "The late Precambrian fossil Kimberella is a mollusc-like bilaterian organism". Nature. 388 (6645): 868–871. Bibcode:1997Natur.388..868F. doi:10.1038/42242. ISSN 0028-0836. S2CID 4395089.
- ^ Mooi R, David B (December 1998). "Evolution Within a Bizarre Phylum: Homologies of the First Echinoderms". American Zoologist. 38 (6): 965–974. doi:10.1093/icb/38.6.965. ISSN 1540-7063.
- ^ McMenamin MA (September 2003). Spriggina is a trilobitoid ecdysozoan. Geoscience Horizons Seattle 2003. Abstracts with Programs. Vol. 35. Boulder, Colo.: Geological Society of America. p. 105. OCLC 249088612. Archived from the original on 12 April 2016. Retrieved 24 November 2007. Paper No. 40-2 presented at the Geological Society of America's 2003 Seattle Annual Meeting (2–5 November 2003) on 2 November 2003, at the Washington State Convention Center.
- ^ Lin JP, Gon III SM, Gehling JG, Babcock LE, Zhao YL, Zhang XL, et al. (2006). "A Parvancorina-like arthropod from the Cambrian of South China". Historical Biology: An International Journal of Paleobiology. 18 (1): 33–45. doi:10.1080/08912960500508689. ISSN 1029-2381. S2CID 85821717.
- ^ Butterfield NJ (December 2006). "Hooking some stem-group "worms": fossil lophotrochozoans in the Burgess Shale". BioEssays. 28 (12): 1161–6. doi:10.1002/bies.20507. PMID 17120226. S2CID 29130876.
- ^ a b Bengtson S (November 2004). "Early skeletal fossils". Paleontological Society Papers. 10: 67–78. doi:10.1017/S1089332600002345.
- ^ Valentine JW (2004). On the Origin of Phyla. Chicago: University Of Chicago Press. p. 7. ISBN 978-0-226-84548-7.
Classifications of organisms in hierarchical systems were in use by the seventeenth and eighteenth centuries. Usually organisms were grouped according to their morphological similarities as perceived by those early workers, and those groups were then grouped according to their similarities, and so on, to form a hierarchy.
- ^ a b Valentine JW (18 June 2004). On the Origin of Phyla. University of Chicago Press. ISBN 9780226845487.
- ^ "WoRMS - World Register of Marine Species". www.marinespecies.org.
- ^ Novak BJ, Fraser D, Maloney TH (February 2020). "Transforming Ocean Conservation: Applying the Genetic Rescue Toolkit". Genes. 11 (2): 209. doi:10.3390/genes11020209. PMC 7074136. PMID 32085502.
- ^ Gould SJ (1990). Wonderful Life: The Burgess Shale and the Nature of History. W. W. Norton. ISBN 978-0-393-30700-9.
- ^ Erwin D, Valentine J, Jablonski D (1997). "Recent fossil finds and new insights into animal development are providing fresh perspectives on the riddle of the explosion of animals during the Early Cambrian". American Scientist (March–April).
- ^ a b Budd GE, Jensen S (May 2000). "A critical reappraisal of the fossil record of the bilaterian phyla". Biological Reviews of the Cambridge Philosophical Society. 75 (2): 253–95. doi:10.1111/j.1469-185X.1999.tb00046.x. PMID 10881389. S2CID 39772232.
- ^ Gould SJ (1989). Wonderful life: the Burgess Shale and the nature of history (First ed.). New York. ISBN 978-0-393-02705-1.
{{cite book}}: CS1 maint: location missing publisher (link) - ^ Budd GE (February 2003). "The Cambrian fossil record and the origin of the phyla". Integrative and Comparative Biology. 43 (1): 157–65. doi:10.1093/icb/43.1.157. PMID 21680420.
- ^ Budd GE (March 1996). "The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group". Lethaia. 29 (1): 1–14. doi:10.1111/j.1502-3931.1996.tb01831.x. ISSN 0024-1164.
- ^ Marshall CR (May 2006). "Explaining the Cambrian 'Explosion' of Animals". Annual Review of Earth and Planetary Sciences. 34: 355–384. Bibcode:2006AREPS..34..355M. doi:10.1146/annurev.earth.33.031504.103001. ISSN 1545-4495. S2CID 85623607.
- ^ King N, Rokas A (October 2017). "Embracing Uncertainty in Reconstructing Early Animal Evolution". Current Biology. 27 (19): R1081–R1088. doi:10.1016/j.cub.2017.08.054. PMC 5679448. PMID 29017048.
- ^ Feuda R, Dohrmann M, Pett W, Philippe H, Rota-Stabelli O, Lartillot N, et al. (December 2017). "Improved Modeling of Compositional Heterogeneity Supports Sponges as Sister to All Other Animals". Current Biology. 27 (24): 3864–3870.e4. doi:10.1016/j.cub.2017.11.008. hdl:11572/302898. PMID 29199080.
- ^ Nielsen C (July 2019). "Early animal evolution: a morphologist's view". Royal Society Open Science. 6 (7): 190638. Bibcode:2019RSOS....690638N. doi:10.1098/rsos.190638. PMC 6689584. PMID 31417759.
- ^ "Porifera (n.)". Online Etymology Dictionary. Retrieved 18 August 2016.
- ^ a b Petralia RS, Mattson MP, Yao PJ (July 2014). "Aging and longevity in the simplest animals and the quest for immortality". Ageing Research Reviews. 16: 66–82. doi:10.1016/j.arr.2014.05.003. PMC 4133289. PMID 24910306.
- ^ Jochum KP, Wang X, Vennemann TW, Sinha B, Müller WE (2012). "Siliceous deep-sea sponge Monorhaphis chuni: A potential paleoclimate archive in ancient animals". Chemical Geology. 300: 143–151. Bibcode:2012ChGeo.300..143J. doi:10.1016/j.chemgeo.2012.01.009.
- ^ Vacelet J, Duport E (November 2004). "Prey capture and digestion in the carnivorous sponge Asbestopluma hypogea (Porifera: Demospongiae)". Zoomorphology. 123 (4): 179–90. doi:10.1007/s00435-004-0100-0. S2CID 24484610.
- ^ "Spongia Linnaeus, 1759". World Register of Marine Species. Retrieved 18 July 2012.
- ^ Rowland SM, Stephens T (2001). "Archaeocyatha: A history of phylogenetic interpretation". Journal of Paleontology. 75 (6): 1065–1078. doi:10.1666/0022-3360(2001)075<1065:AAHOPI>2.0.CO;2. JSTOR 1307076. S2CID 86211946.
- ^ Sperling EA, Pisani D, Peterson KJ (1 January 2007). "Poriferan paraphyly and its implications for Precambrian palaeobiology" (PDF). Geological Society, London, Special Publications. 286 (1): 355–368. Bibcode:2007GSLSP.286..355S. doi:10.1144/SP286.25. S2CID 34175521. Archived from the original (PDF) on 9 May 2009. Retrieved 22 August 2012.
- ^ Ruppert EE, Fox RS, Barnes RD (2004). Invertebrate Zoology (7 ed.). Brooks / Cole. pp. 182–195. ISBN 978-0-03-025982-1.
- ^ Mills CE. "Ctenophores – some notes from an expert". Retrieved 5 February 2009.
- ^ a b Brusca RC, Brusca GJ (2003). Invertebrates (Second ed.). Sinauer Associates. ISBN 978-0-87893-097-5.
- ^ Le Page M (March 2019). "Animal with an anus that comes and goes could reveal how ours evolved". New Scientist.
- ^ Martindale MQ, Finnerty JR, Henry JQ (September 2002). "The Radiata and the evolutionary origins of the bilaterian body plan". Molecular Phylogenetics and Evolution. 24 (3): 358–65. doi:10.1016/s1055-7903(02)00208-7. PMID 12220977.
- ^ Placozoa at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- ^ Wehner R, Gehring W (June 2007). Zoologie (in German) (24th ed.). Stuttgart: Thieme. p. 696.
- ^ Schulze FE (1883). "Trichoplax adhaerens n. g., n. s.". Zoologischer Anzeiger. Vol. 6. Amsterdam and Jena: Elsevier. p. 92.
- ^ Eitel M, Francis WR, Osigus HJ, Krebs S, Vargas S, Blum H, et al. (13 October 2017). "A taxogenomics approach uncovers a new genus in the phylum Placozoa". bioRxiv: 202119. doi:10.1101/202119. S2CID 89829846.
- ^ Osigus HJ, Rolfes S, Herzog R, Kamm K, Schierwater B (March 2019). "Polyplacotoma mediterranea is a new ramified placozoan species". Current Biology. 29 (5): R148–R149. doi:10.1016/j.cub.2019.01.068. PMID 30836080.
- ^ "Trichoplax adhaerens". WoRMS. 2009.
- ^ Smith CL, Varoqueaux F, Kittelmann M, Azzam RN, Cooper B, Winters CA, et al. (July 2014). "Novel cell types, neurosecretory cells, and body plan of the early-diverging metazoan Trichoplax adhaerens". Current Biology. 24 (14): 1565–1572. doi:10.1016/j.cub.2014.05.046. PMC 4128346. PMID 24954051.
- ^ Barnes RD (1982). Invertebrate Zoology. Philadelphia: Holt-Saunders International. pp. 84–85. ISBN 978-0-03-056747-6.
- ^ Zhang ZQ (2011). "Animal biodiversity: An introduction to higher-level classification and taxonomic richness" (PDF). Zootaxa. 3148: 7–12. doi:10.11646/zootaxa.3148.1.3.
- ^ "Nematostella vectensis v1.0". Genome Portal. University of California. Retrieved 19 January 2014.
- ^ "Nematostella". Nematostella.org. Archived from the original on 8 May 2006. Retrieved 18 January 2014.
- ^ a b Genikhovich G, Technau U (September 2009). "The starlet sea anemone Nematostella vectensis: an anthozoan model organism for studies in comparative genomics and functional evolutionary developmental biology". Cold Spring Harbor Protocols. 2009 (9): pdb.emo129. doi:10.1101/pdb.emo129. PMID 20147257.
- ^ "Where Does Our Head Come From? Brainless Sea Anemone Sheds New Light on the Evolutionary Origin of the Head". Science Daily. 12 February 2013. Retrieved 18 January 2014.
- ^ Sinigaglia C, Busengdal H, Leclère L, Technau U, Rentzsch F (2013). "The bilaterian head patterning gene six3/6 controls aboral domain development in a cnidarian". PLOS Biology. 11 (2): e1001488. doi:10.1371/journal.pbio.1001488. PMC 3586664. PMID 23483856.
- ^ "Red Paper Lantern Jellyfish". Real Monstrosities. Retrieved 25 October 2015.
- ^ "Blue Buttons in Florida". BeachHunter.net.
- ^ Karleskint G, Turner R, Small J (2012). Introduction to Marine Biology (4th ed.). Cengage Learning. p. 445. ISBN 978-1-133-36446-7.
- ^ Bavestrello G, Sommer C, Sarà M (1992). "Bi-directional conversion in Turritopsis nutricula (Hydrozoa)". Scientia Marina. 56 (2–3): 137–140.
- ^ Piraino S, Boero F, Aeschbach B, Schmid V (1996). "Reversing the life cycle: medusae transforming into polyps and cell transdifferentiation in Turritopsis nutricula (Cnidaria, Hydrozoa)". Biological Bulletin. 190 (3): 302–312. doi:10.2307/1543022. JSTOR 1543022. PMID 29227703. S2CID 3956265.
- ^ Fenner PJ, Williamson JA (1996). "Worldwide deaths and severe envenomation from jellyfish stings". The Medical Journal of Australia. 165 (11–12): 658–61. doi:10.5694/j.1326-5377.1996.tb138679.x. PMID 8985452. S2CID 45032896.
- ^ a b c Cannon JT, Vellutini BC, Smith J, Ronquist F, Jondelius U, Hejnol A (February 2016). "Xenacoelomorpha is the sister group to Nephrozoa". Nature. 530 (7588): 89–93. Bibcode:2016Natur.530...89C. doi:10.1038/nature16520. PMID 26842059. S2CID 205247296.
- ^ a b Minelli A (2009). Perspectives in Animal Phylogeny and Evolution. Oxford University Press. p. 53. ISBN 978-0-19-856620-5.
- ^ a b c Brusca RC (2016). Introduction to the Bilateria and the Phylum Xenacoelomorpha | Triploblasty and Bilateral Symmetry Provide New Avenues for Animal Radiation (PDF). Sinauer Associates. pp. 345–372. ISBN 978-1605353753.
{{cite book}}:|work=ignored (help) - ^ Finnerty JR (November 2005). "Did internal transport, rather than directed locomotion, favor the evolution of bilateral symmetry in animals?" (PDF). BioEssays. 27 (11): 1174–80. doi:10.1002/bies.20299. PMID 16237677. Archived from the original (PDF) on 10 August 2014. Retrieved 27 August 2019.
- ^ Quillin KJ (May 1998). "Ontogenetic scaling of hydrostatic skeletons: geometric, static stress and dynamic stress scaling of the earthworm lumbricus terrestris". The Journal of Experimental Biology. 201 (12): 1871–83. doi:10.1242/jeb.201.12.1871. PMID 9600869.
- ^ Specktor B (26 March 2020). "This primeval worm may be the ancestor of all animals]". Live Science'.
- ^ a b Wade N (30 January 2017). "This Prehistoric Human Ancestor Was All Mouth". The New York Times. Retrieved 31 January 2017.
- ^ a b Han J, Morris SC, Ou Q, Shu D, Huang H (February 2017). "Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China)". Nature. 542 (7640): 228–231. Bibcode:2017Natur.542..228H. doi:10.1038/nature21072. PMID 28135722. S2CID 353780.
- ^ "Cornwall – Nature – Superstar Worm". BBC.
- ^ Mark Carwardine (1995) The Guinness Book of Animal Records. Guinness Publishing. p. 232.
- ^ "The Persistent Parasites". Time. 8 April 1957. Archived from the original on 27 June 2008.
- ^ Hargis W, ed. (1985). Parasitology and pathology of marine organisms of the world ocean (Technical report). National Oceanic and Atmospheric Administration.
- ^ "Classification of Animal Parasites". Archived from the original on 14 September 2006.
- ^ Garcia LS (October 1999). "Classification of human parasites, vectors, and similar organisms". Clinical Infectious Diseases. 29 (4): 734–6. doi:10.1086/520425. PMID 10589879.
- ^ Hodda M (2011). "Phylum Nematoda Cobb, 1932. In: Zhang, Z.-Q. Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness". Zootaxa. 3148: 63–95. doi:10.11646/zootaxa.3148.1.11.
- ^ Zhang Z (2013). "Animal biodiversity: An update of classification and diversity in 2013. In: Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013)". Zootaxa. 3703 (1): 5–11. doi:10.11646/zootaxa.3703.1.3. S2CID 85252974.
- ^ Lambshead PJ (1993). "Recent developments in marine benthic biodiversity research". Oceanis. 19 (6): 5–24.
- ^ Borgonie G, García-Moyano A, Litthauer D, Bert W, Bester A, van Heerden E, et al. (June 2011). "Nematoda from the terrestrial deep subsurface of South Africa". Nature. 474 (7349): 79–82. Bibcode:2011Natur.474...79B. doi:10.1038/nature09974. hdl:1854/LU-1269676. PMID 21637257. S2CID 4399763.
- ^ Danovaro R, Gambi C, Dell'Anno A, Corinaldesi C, Fraschetti S, Vanreusel A, et al. (January 2008). "Exponential decline of deep-sea ecosystem functioning linked to benthic biodiversity loss". Current Biology. 18 (1): 1–8. doi:10.1016/j.cub.2007.11.056. PMID 18164201. S2CID 15272791.
- "Deep-sea species' loss could lead to oceans' collapse, study suggests". EurekAlert! (Press release). 27 December 2007.
- ^ Platt HM (1994). "foreword". In Lorenzen S, Lorenzen SA (eds.). The phylogenetic systematics of freeliving nematodes. London: The Ray Society. ISBN 978-0-903874-22-9.
- ^ Barnes RS, Calow P, Olive PJ (2001). The Invertebrates, A Synthesis (3rd ed.). UK: Blackwell Science.
- ^ "Tyrian Purple". Green Lion. 28 February 2014. Archived from the original on 28 February 2014.
- ^ Chapman AD (2009). Numbers of Living Species in Australia and the World (2nd ed.). Canberra: Australian Biological Resources Study. ISBN 978-0-642-56860-1.
- ^ Hancock R (2008). "Recognising research on molluscs". Australian Museum. Archived from the original on 30 May 2009. Retrieved 9 March 2009.
- ^ Ponder WF, Lindberg DR, eds. (2008). Phylogeny and Evolution of the Mollusca. Berkeley: University of California Press. p. 481. ISBN 978-0-520-25092-5.
- ^ Munro D, Blier PU (October 2012). "The extreme longevity of Arctica islandica is associated with increased peroxidation resistance in mitochondrial membranes". Aging Cell. 11 (5): 845–55. doi:10.1111/j.1474-9726.2012.00847.x. PMID 22708840. S2CID 205634828.
- ^ "Welcome to CephBase". CephBase. Archived from the original on 12 January 2016. Retrieved 29 January 2016.
- ^ Wilbur KM, Clarke, M.R., Trueman, E.R., eds. (1985), The Mollusca, vol. 12. Paleontology and neontology of Cephalopods, New York: Academic Press, ISBN 0-12-728702-7
- ^ "Are there any freshwater cephalopods?". Australian Broadcasting Corporation. 16 January 2013.
- ^ Callaway E (2 June 2008). "Simple-Minded Nautilus Shows Flash of Memory". New Scientist. Retrieved 7 March 2012.
- ^ Phillips K (15 June 2008). "Living Fossil Memories". Journal of Experimental Biology. 211 (12): iii. doi:10.1242/jeb.020370. S2CID 84279320.
- ^ Crook R, Basil J (2008). "A biphasic memory curve in the chambered nautilus, Nautilus pompilius L. (Cephalopoda: Nautiloidea)". Journal of Experimental Biology. 211 (12): 1992–1998. doi:10.1242/jeb.018531. PMID 18515730. S2CID 6305526.
- ^ Black R (26 April 2008). "Colossal squid out of the freezer". BBC News.
- ^ a b Ruppert RE, Fox RS, Barnes RD (2004). Invertebrate Zoology (7th ed.). Cengage Learning. ISBN 978-81-315-0104-7.
- ^ Hayward PJ (1996). Handbook of the Marine Fauna of North-West Europe. Oxford University Press. ISBN 978-0-19-854055-7.
- ^ Wilson HM, Anderson LI (January 2004). "Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland". Journal of Paleontology. 78 (1): 169–184. doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2. S2CID 131201588.
- ^ Suarez SE, Brookfield ME, Catlos EJ, Stöckli DF (2017). "A U-Pb zircon age constraint on the oldest-recorded air-breathing land animal". PLOS ONE. 12 (6): e0179262. Bibcode:2017PLoSO..1279262S. doi:10.1371/journal.pone.0179262. PMC 5489152. PMID 28658320.
- ^ Campbell LI, Rota-Stabelli O, Edgecombe GD, Marchioro T, Longhorn SJ, Telford MJ, et al. (September 2011). "MicroRNAs and phylogenomics resolve the relationships of Tardigrada and suggest that velvet worms are the sister group of Arthropoda". Proceedings of the National Academy of Sciences of the United States of America. 108 (38): 15920–4. Bibcode:2011PNAS..10815920C. doi:10.1073/pnas.1105499108. PMC 3179045. PMID 21896763.
- ^ Smith FW, Goldstein B (May 2017). "Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda". Arthropod Structure & Development. 46 (3): 328–340. doi:10.1016/j.asd.2016.10.005. PMID 27725256.
- ^ Budd GE (2001). "Why are arthropods segmented?". Evolution & Development. 3 (5): 332–42. doi:10.1046/j.1525-142X.2001.01041.x. PMID 11710765. S2CID 37935884.
- ^ "David Attenborough's First Life". Archived from the original on 26 January 2011. Retrieved 10 March 2011.
- ^ Braddy SJ, Poschmann M, Tetlie OE (February 2008). "Giant claw reveals the largest ever arthropod". Biology Letters. 4 (1): 106–9. doi:10.1098/rsbl.2007.0491. PMC 2412931. PMID 18029297.
- ^ Daniel C (21 November 2007). "Giant sea scorpion discovered". Nature. doi:10.1038/news.2007.272. Retrieved 10 June 2013.
- ^ Bicknell, Russell D. C.; Pates, Stephen (2020). "Pictorial Atlas of Fossil and Extant Horseshoe Crabs, With Focus on Xiphosurida". Frontiers in Earth Science. 8: 98. Bibcode:2020FrEaS...8...98B. doi:10.3389/feart.2020.00098. ISSN 2296-6463.
- ^ "An ugly giant crab of Japan". Popular Science. 96 (6): 42. 1920.
- ^ Currie DR, Ward TM (2009). South Australian Giant Crab (Pseudocarcinus gigas) Fishery (PDF). South Australian Research and Development Institute. Fishery Assessment Report for PIRSA. Archived from the original (PDF) on 28 March 2012. Retrieved 9 December 2013.
- ^ Kilday P (28 September 2005). "Mantis shrimp boasts most advanced eyes". The Daily Californian. Archived from the original on 29 September 2012. Retrieved 23 September 2016.
- ^ Patek SN, Caldwell RL (2005). "Extreme impact and cavitation forces of a biological hammer: strike forces of the peacock mantis shrimp". Journal of Experimental Biology. 208 (19): 3655–3664. doi:10.1242/jeb.01831. PMID 16169943. S2CID 312009.
- ^ Han, Jian; Morris, Simon Conway; Ou, Qiang; Shu, Degan; Huang, Hai (2017). "Meiofaunal deuterostomes from the basal Cambrian of Shaanxi (China)". Nature. 542 (7640): 228–231. Bibcode:2017Natur.542..228H. doi:10.1038/nature21072. ISSN 1476-4687. PMID 28135722. S2CID 353780.
- ^ Liu, Yunhuan; Carlisle, Emily; Zhang, Huaqiao; Yang, Ben; Steiner, Michael; Shao, Tiequan; Duan, Baichuan; Marone, Federica; Xiao, Shuhai; Donoghue, Philip C. J. (17 August 2022). "Saccorhytus is an early ecdysozoan and not the earliest deuterostome". Nature. 609 (7927): 541–546. Bibcode:2022Natur.609..541L. doi:10.1038/s41586-022-05107-z. hdl:1983/454e7bec-4cd4-4121-933e-abeab69e96c1. ISSN 1476-4687. PMID 35978194. S2CID 251646316.
- ^ "Animal Diversity Web - Echinodermata". University of Michigan Museum of Zoology. Retrieved 26 August 2012.
- ^ "Echinoderm | Definition, Characteristics, Species, & Facts | Britannica". www.britannica.com. 8 June 2023. Retrieved 24 June 2023.
- ^ Fox R. "Asterias forbesi". Invertebrate Anatomy OnLine. Lander University. Retrieved 14 June 2014.
- ^ Holsinger, K. (2005). Keystone species. Retrieved 10 May 2010, from Holsinger K (11 October 2005). "Keystone species". University of Connecticut. Archived from the original on 30 June 2010. Retrieved 12 May 2010.
- ^ Simakov O, Kawashima T, Marlétaz F, Jenkins J, Koyanagi R, Mitros T, et al. (November 2015). "Hemichordate genomes and deuterostome origins". Nature. 527 (7579): 459–65. Bibcode:2015Natur.527..459S. doi:10.1038/nature16150. PMC 4729200. PMID 26580012.
- ^ "How humans got a pharynx from this 'ugly beast'". Futurity. 23 November 2015.
- ^ a b c d Clark MA, Choi J, Douglas M (2018). "Chordates". Biology (2nd ed.). OpenStax. ISBN 978-1-947172-95-1.. Modified text was copied from this source, which is available under a Creative Commons Attribution 4.0 International License
- ^ "The secret to an Oesia life: Prehistoric worm built tube-like 'houses' on sea floor". phys.org.
- ^ Barnes RD (1982). Invertebrate Zoology. Philadelphia, PA: Holt-Saunders International. pp. 1018–1026. ISBN 978-0-03-056747-6.
- ^ "Secondary organizers of the early brain and the location of the meso-diencephalic dopaminergic precursor cells". Life Map. Archived from the original on 10 March 2014. Retrieved 10 March 2014.
- ^ Pani AM, Mullarkey EE, Aronowicz J, Assimacopoulos S, Grove EA, Lowe CJ (March 2012). "Ancient deuterostome origins of vertebrate brain signalling centres". Nature. 483 (7389). ScienceLife: 289–94. Bibcode:2012Natur.483..289P. doi:10.1038/nature10838. PMC 3719855. PMID 22422262.
- ^ "Chordates". OpenStax. 9 May 2019.
- ^ Gewin V (2005). "Functional genomics thickens the biological plot". PLOS Biology. 3 (6): e219. doi:10.1371/journal.pbio.0030219. PMC 1149496. PMID 15941356.
- ^ Timmer J (19 June 2008). "Lancelet (amphioxus) genome and the origin of vertebrates". Ars Technica.
- ^ Lemaire P (2011). "Evolutionary crossroads in developmental biology: the tunicates". Development. 138 (11): 2143–2152. doi:10.1242/dev.048975. PMID 21558365. S2CID 40452112.
- ^ "FishBase: A Global Information System on Fishes". FishBase. Retrieved 17 January 2017.
- ^ "How Many Fish In The Sea? Census Of Marine Life Launches First Report". Science Daily. Retrieved 17 January 2017.
- ^ Docker MF (November 2006). "Bill Beamish's Contributions to Lamprey Research and Recent Advances in the Field". Guelph Ichthyology Reviews. 7.
- ^ Hardisty MW, Potter IC (1971). Hardisty MW, Potter IC (eds.). The Biology of Lampreys (1st ed.). Academic Press. ISBN 978-0-123-24801-5.
- ^ Gill HS, Renaud CB, Chapleau F, Mayden RL, Potter IC (2003). "Phylogeny of Living Parasitic Lampreys (Petromyzontiformes) Based on Morphological Data". Copeia. 2003 (4): 687–703. doi:10.1643/IA02-085.1. S2CID 85969032.
- ^ "Myxini". University of California Museum of Paleontology. Archived from the original on 15 December 2017. Retrieved 17 January 2017.
- ^ Green SA, Bronner ME (2014). "The lamprey: a jawless vertebrate model system for examining origin of the neural crest and other vertebrate traits". Differentiation; Research in Biological Diversity. 87 (1–2): 44–51. doi:10.1016/j.diff.2014.02.001. PMC 3995830. PMID 24560767.
- ^ Stock DW, Whitt GS (August 1992). "Evidence from 18S ribosomal RNA sequences that lampreys and hagfishes form a natural group". Science. 257 (5071): 787–9. Bibcode:1992Sci...257..787S. doi:10.1126/science.1496398. PMID 1496398.
- ^ Nicholls H (September 2009). "Evolution: Mouth to mouth". Nature. 461 (7261): 164–6. doi:10.1038/461164a. PMID 19741680.
- ^ McCoy VE, Saupe EE, Lamsdell JC, Tarhan LG, McMahon S, Lidgard S, et al. (April 2016). "The 'Tully monster' is a vertebrate". Nature. 532 (7600): 496–9. Bibcode:2016Natur.532..496M. doi:10.1038/nature16992. PMID 26982721. S2CID 205247805.
- ^ Sallan L, Giles S, Sansom RS, Clarke JT, Johanson Z, Sansom IJ, et al. (20 February 2017). "The 'Tully Monster' is not a vertebrate: characters, convergence and taphonomy in Palaeozoic problematic animals" (PDF). Palaeontology. 60 (2): 149–157. Bibcode:2017Palgy..60..149S. doi:10.1111/pala.12282.
- ^ Geggel L (4 May 2020). "Ancient 'Tully monster' was a vertebrate, not a spineless blob, study claims". Live Science.
- ^ McCoy VE, Wiemann J, Lamsdell JC, Whalen CD, Lidgard S, Mayer P, et al. (September 2020). "Chemical signatures of soft tissues distinguish between vertebrates and invertebrates from the Carboniferous Mazon Creek Lagerstätte of Illinois". Geobiology. 18 (5): 560–565. Bibcode:2020Gbio...18..560M. doi:10.1111/gbi.12397. PMID 32347003. S2CID 216646333.
- ^ Mikami, Tomoyuki; Ikeda, Takafumi; Muramiya, Yusuke; Hirasawa, Tatsuya; Iwasaki, Wataru (2023). Cherns, Lesley (ed.). "Three-dimensional anatomy of the Tully monster casts doubt on its presumed vertebrate affinities". Palaeontology. 66 (2). doi:10.1111/pala.12646. ISSN 0031-0239. S2CID 258198566.
- ^ Kimmel CB, Miller CT, Keynes RJ (2001). "Neural crest patterning and the evolution of the jaw". Journal of Anatomy. 199 (Pt 1-2): 105–20. doi:10.1017/S0021878201008068. PMC 1594948. PMID 11523812.
- ^ Gai Z, Zhu M (2012). "The origin of the vertebrate jaw: Intersection between developmental biology-based model and fossil evidence". Chinese Science Bulletin. 57 (30): 3819–3828. Bibcode:2012ChSBu..57.3819G. doi:10.1007/s11434-012-5372-z.
- ^ Maisey JG (2000). Discovering Fossil Fishes. Westview Press. pp. 1–223. ISBN 978-0-8133-3807-1.
- ^ a b Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, et al. (2008). "Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?" (PDF). Journal of Zoology. 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x.
- ^ Pimiento C, Ehret DJ, Macfadden BJ, Hubbell G (May 2010). Stepanova A (ed.). "Ancient nursery area for the extinct giant shark megalodon from the Miocene of Panama". PLOS ONE. 5 (5): e10552. Bibcode:2010PLoSO...510552P. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893.
- ^ Lambert O, Bianucci G, Post K, de Muizon C, Salas-Gismondi R, Urbina M, Reumer J (July 2010). "The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru". Nature. 466 (7302): 105–8. Bibcode:2010Natur.466..105L. doi:10.1038/nature09067. PMID 20596020. S2CID 4369352.
- ^ Nielsen J, Hedeholm RB, Heinemeier J, Bushnell PG, Christiansen JS, Olsen J, et al. (August 2016). "Eye lens radiocarbon reveals centuries of longevity in the Greenland shark (Somniosus microcephalus)". Science. 353 (6300): 702–4. Bibcode:2016Sci...353..702N. doi:10.1126/science.aaf1703. hdl:2022/26597. PMID 27516602. S2CID 206647043.
- Enrico de Lazaro (12 August 2016). "Greenland Sharks are Longest-Lived Vertebrates on Earth, Marine Biologists Say". Science News.
- ^ Marshall A, Bennett MB, Kodja G, Hinojosa-Alvarez S, Galvan-Magana F, Harding M, et al. (2011). "Manta birostris". IUCN Red List of Threatened Species. 2011: e.T198921A9108067. doi:10.2305/IUCN.UK.2011-2.RLTS.T198921A9108067.en.
- ^ Black R (11 June 2007). "Sawfish protection acquires teeth". BBC News.
- ^ Near TJ, Eytan RI, Dornburg A, Kuhn KL, Moore JA, Davis MP, et al. (August 2012). "Resolution of ray-finned fish phylogeny and timing of diversification". Proceedings of the National Academy of Sciences of the United States of America. 109 (34): 13698–703. Bibcode:2012PNAS..10913698N. doi:10.1073/pnas.1206625109. PMC 3427055. PMID 22869754.
- ^ a b Zhu M, Zhao W, Jia L, Lu J, Qiao T, Qu Q (March 2009). "The oldest articulated osteichthyan reveals mosaic gnathostome characters". Nature. 458 (7237): 469–74. Bibcode:2009Natur.458..469Z. doi:10.1038/nature07855. PMID 19325627. S2CID 669711.
- ^ Clack, J. A. (2002) Gaining Ground. Indiana University
- ^ "Chondrosteans: Sturgeon Relatives". paleos.com. Archived from the original on 25 December 2010.
- ^ López-Arbarello A (2012). "Phylogenetic interrelationships of ginglymodian fishes (Actinopterygii: Neopterygii)". PLOS ONE. 7 (7): e39370. Bibcode:2012PLoSO...739370L. doi:10.1371/journal.pone.0039370. PMC 3394768. PMID 22808031.
- ^ Berra TM (2008). Freshwater Fish Distribution. University of Chicago Press. p. 55. ISBN 978-0-226-04443-9.
- ^ Lackmann AR, Andrews AH, Butler MG, Bielak-Lackmann ES, Clark ME (23 May 2019). "Bigmouth Buffalo Ictiobus cyprinellus sets freshwater teleost record as improved age analysis reveals centenarian longevity". Communications Biology. 2 (1): 197. doi:10.1038/s42003-019-0452-0. PMC 6533251. PMID 31149641.
- ^ a b Benton M (2005). "The Evolution of Fishes After the Devonian". Vertebrate Palaeontology (3rd ed.). John Wiley & Sons. pp. 175–84. ISBN 978-1-4051-4449-0.
- ^ Bone Q, Moore R (2008). Biology of Fishes. Garland Science. p. 29. ISBN 978-0-415-37562-7.
- ^ Dorit R, Walker WF, Barnes RD (1991). Zoology. Saunders College Publishing. pp. 67–69. ISBN 978-0-03-030504-7.
- ^ "Scientists Describe the World's Smallest, Lightest Fish". Scripps Institution of Oceanography. 20 July 2004. Archived from the original on 5 March 2016. Retrieved 9 April 2016.
- ^ Roach J (13 May 2003). "World's Heaviest Bony Fish Discovered?". National Geographic News. Archived from the original on 17 May 2003. Retrieved 9 January 2016.
- ^ "Summary Statistics for Globally Threatened Species" (PDF). IUCN Red List of Threatened Species. The World Conservation Union. Autumn 2014.
Table 1: Numbers of threatened species by major groups of organisms (1996–2014)
- ^ Narkiewicz K, Narkiewicz M (January 2015). "The age of the oldest tetrapod tracks from Zachełmie, Poland". Lethaia. 48 (1): 10–12. doi:10.1111/let.12083. ISSN 0024-1164.
- ^ Long JA, Gordon MS (September–October 2004). "The greatest step in vertebrate history: a paleobiological review of the fish-tetrapod transition" (PDF). Physiological and Biochemical Zoology. 77 (5): 700–19. doi:10.1086/425183. PMID 15547790. S2CID 1260442.
- ^ Shubin N (2008). Your Inner Fish: A Journey Into the 3.5-Billion-Year History of the Human Body. New York: Pantheon Books. ISBN 978-0-375-42447-2.
- ^ Laurin M (2010). How Vertebrates Left the Water. Berkeley, California, USA.: University of California Press. ISBN 978-0-520-26647-6.
- ^ Canoville A, Laurin M (2010). "Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on paleobiological inferences". Biological Journal of the Linnean Society. 100 (2): 384–406. doi:10.1111/j.1095-8312.2010.01431.x.
- ^ Laurin M, Canoville A, Quilhac A (August 2009). "Use of paleontological and molecular data in supertrees for comparative studies: the example of lissamphibian femoral microanatomy". Journal of Anatomy. 215 (2): 110–23. doi:10.1111/j.1469-7580.2009.01104.x. PMC 2740958. PMID 19508493.
- ^ Hopkins GR, Brodie Jr ED (2015). "Occurrence of Amphibians in Saline Habitats: A Review and Evolutionary Perspective". Herpetological Monographs. 29 (1): 1–27. doi:10.1655/HERPMONOGRAPHS-D-14-00006. S2CID 83659304.
- ^ Natchev N, Tzankov N, Geme R (2011). "Green frog invasion in the Black Sea: habitat ecology of the Pelophylax esculentus complex (Anura, Amphibia) population in the region of Shablenska Tuzla lagoon in Bulgaria" (PDF). Herpetology Notes. 4: 347–351. Archived from the original (PDF) on 24 September 2015. Retrieved 11 August 2016.
- ^ Sander PM (August 2012). "Paleontology. Reproduction in early amniotes". Science. 337 (6096): 806–8. Bibcode:2012Sci...337..806S. doi:10.1126/science.1224301. PMID 22904001. S2CID 7041966.
- ^ Modesto SP, Anderson JS (October 2004). "The phylogenetic definition of reptilia". Systematic Biology. 53 (5): 815–21. doi:10.1080/10635150490503026. PMID 15545258.

- ^ Gauthier JA, Kluge AG, Rowe T (1988). "The early evolution of the Amniota". In Benton MJ (ed.). The Phylogeny and Classification of the Tetrapods. Vol. 1. Oxford: Clarendon Press. pp. 103–155. ISBN 978-0-19-857705-8.
- ^ Laurin M, Reisz RR (1995). "A reevaluation of early amniote phylogeny" (PDF). Zoological Journal of the Linnean Society. 113 (2): 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x. Archived from the original (PDF) on 8 June 2019. Retrieved 14 August 2016.
- ^ Modesto SP (1999). "Observations of the structure of the Early Permian reptile Stereosternum tumidum Cope". Palaeontologia Africana. 35: 7–19.
- ^ Rasmussen AR, Murphy JC, Ompi M, Gibbons JW, Uetz P (8 November 2011). "Marine reptiles". PLOS ONE. 6 (11): e27373. Bibcode:2011PLoSO...627373R. doi:10.1371/journal.pone.0027373. PMC 3210815. PMID 22087300.
- ^ Stidworthy J (1974). Snakes of the World. Grosset & Dunlap Inc. p. 160. ISBN 978-0-448-11856-7.
- ^ "Sea snakes" (PDF). Food and Agriculture Organization of the United Nations. Retrieved 22 August 2020.
- ^ Rasmussen AR, Murphy JC, Ompi M, Gibbons JW, Uetz P (2011). "Marine reptiles". PLOS ONE. 6 (11): e27373. Bibcode:2011PLoSO...627373R. doi:10.1371/journal.pone.0027373. PMC 3210815. PMID 22087300.
- ^ Martill D.M. (1993). "Soupy Substrates: A Medium for the Exceptional Preservation of Ichthyosaurs of the Posidonia Shale (Lower Jurassic) of Germany". Kaupia - Darmstädter Beiträge zur Naturgeschichte, 2 : 77-97.
- ^ Gould SJ (1993). "Bent Out of Shape". Eight Little Piggies: Reflections in Natural History. Norton. pp. 179–94. ISBN 978-0-393-31139-6.
- ^ "Sardine Run Shark Feeding Frenzy Phenomenon in Africa". Archived from the original on 2 December 2008.
- ^ "The Society for Marine Mammalogy's Taxonomy Committee List of Species and subspecies". Society for Marine Mammalogy. October 2015. Archived from the original on 6 January 2015. Retrieved 23 November 2015.
- ^ Romer AS, Parsons TS (1986). The Vertebrate Body. Sanders College Publishing. p. 96. ISBN 978-0-03-058446-6.
- ^ "Blue whale". World Wide Fund For Nature. Retrieved 15 August 2016.
- ^ Marino L (2004). "Cetacean Brain Evolution: Multiplication Generates Complexity" (PDF). International Society for Comparative Psychology (17): 1–16. Archived from the original (PDF) on 16 September 2018. Retrieved 15 August 2016.
- ^ a b Campbell NA, Reece JB, Urry LA, Cain ML, Wasserman SA, Minorsky PV, Jackson RB (2008). Biology (8 ed.). San Francisco: Pearson – Benjamin Cummings. ISBN 978-0-321-54325-7.
- ^ Barrie FR, Buck WR, Demoulin V, Greuter W, Hawksworth DL, Herendeen PS, et al., eds. (2012). International Code of Nomenclature for algae, fungi, and plants (Melbourne Code), Adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011 (electronic ed.). International Association for Plant Taxonomy. Retrieved 14 May 2017.
- ^ Walsh PJ, Smith S, Fleming L, Solo-Gabriele H, Gerwick WH, eds. (2 September 2011). "Cyanobacteria and cyanobacterial toxins". Oceans and Human Health: Risks and Remedies from the Seas. Academic Press. pp. 271–296. ISBN 978-0-08-087782-2.
- ^ "The Rise of Oxygen - Astrobiology Magazine". Astrobiology Magazine. 30 July 2003. Retrieved 6 April 2016.
- ^ Flannery DT, Walter RM (2012). "Archean tufted microbial mats and the Great Oxidation Event: new insights into an ancient problem". Australian Journal of Earth Sciences. 59 (1): 1–11. Bibcode:2012AuJES..59....1F. doi:10.1080/08120099.2011.607849. S2CID 53618061.
- ^ Rothschild L (September 2003). "Understand the evolutionary mechanisms and environmental limits of life". NASA. Archived from the original on 29 March 2012. Retrieved 13 July 2009.
- ^ Nadis S (December 2003). "The cells that rule the seas" (PDF). Scientific American. 289 (6): 52–3. Bibcode:2003SciAm.289f..52N. doi:10.1038/scientificamerican1203-52. PMID 14631732. Archived from the original (PDF) on 19 April 2014. Retrieved 2 June 2019.
- ^ "The Most Important Microbe You've Never Heard Of". npr.org.
- ^ Flombaum P, Gallegos JL, Gordillo RA, Rincón J, Zabala LL, Jiao N, et al. (June 2013). "Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus". Proceedings of the National Academy of Sciences of the United States of America. 110 (24): 9824–9. Bibcode:2013PNAS..110.9824F. doi:10.1073/pnas.1307701110. PMC 3683724. PMID 23703908.
- ^ Nabors MW (2004). Introduction to Botany. San Francisco, CA: Pearson Education, Inc. ISBN 978-0-8053-4416-5.
- ^ Allaby M, ed. (1992). "Algae". The Concise Dictionary of Botany. Oxford: Oxford University Press.
- ^ Guiry MD (October 2012). "How Many Species of Algae Are There?". Journal of Phycology. 48 (5): 1057–63. doi:10.1111/j.1529-8817.2012.01222.x. PMID 27011267. S2CID 30911529.
- ^ a b Guiry MD, Guiry GM (2016). "Algaebase". www.algaebase.org. Retrieved 20 November 2016.
- ^ Thomas D (2002). Seaweeds. Life Series. Natural History Museum, London. ISBN 978-0-565-09175-0.
- ^ Hoek C, Mann D, Jahns HM, Jahns M (1995). Algae: an introduction to phycology. Cambridge University Press. p. 166. ISBN 9780521316873. OCLC 443576944.
- ^ Tréguer P, Nelson DM, Van Bennekom AJ, Demaster DJ, Leynaert A, Quéguiner B (April 1995). "The silica balance in the world ocean: a reestimate". Science. 268 (5209): 375–9. Bibcode:1995Sci...268..375T. doi:10.1126/science.268.5209.375. PMID 17746543. S2CID 5672525.
- ^ "King's College London - Lake Megachad". www.kcl.ac.uk. Retrieved 5 May 2018.
- ^ Gómez F (2012). "A checklist and classification of living dinoflagellates (Dinoflagellata, Alveolata)". CICIMAR Oceánides. 27 (1): 65–140. doi:10.37543/oceanides.v27i1.111.
- ^ Stoecker DK (1999). "Mixotrophy among Dinoflagellates". The Journal of Eukaryotic Microbiology. 46 (4): 397–401. doi:10.1111/j.1550-7408.1999.tb04619.x. S2CID 83885629.
- ^ Starckx S (31 October 2012). "A place in the sun - Algae is the crop of the future, according to researchers in Geel] Flanders Today". Archived from the original on 4 March 2016. Retrieved 8 December 2012.
- ^ Duval B, Margulis L (1995). "The microbial community of Ophrydium versatile colonies: endosymbionts, residents, and tenants". Symbiosis. 18: 181–210. PMID 11539474.
- ^ Wernberg, T., Krumhansl, K., Filbee-Dexter, K. and Pedersen, M.F. (2019) "Status and trends for the world's kelp forests". In: World seas: an environmental evaluation, pages 57–78). Academic Press. doi:10.1016/B978-0-12-805052-1.00003-6.
- ^ Mann KH (December 1973). "Seaweeds: Their Productivity and Strategy for Growth: The role of large marine algae in coastal productivity is far more important than has been suspected". Science. 182 (4116): 975–81. Bibcode:1973Sci...182..975M. doi:10.1126/science.182.4116.975. PMID 17833778. S2CID 26764207.
- ^ Tunnell JW, Chávez EA, Withers K (2007). Coral reefs of the southern Gulf of Mexico. Texas A&M University Press. p. 91. ISBN 978-1-58544-617-9.
- ^ "Caulerpa taxifolia (killer algae)". Invasive Species Compendium. Centre for Agriculture and Bioscience International. 6 November 2018.
- ^ Mandoli DF (June 1998). "Elaboration of Body Plan and Phase Change During Development of Acetabularia: How Is the Complex Architecture of a Giant Unicell Built?". Annual Review of Plant Physiology and Plant Molecular Biology. 49: 173–198. doi:10.1146/annurev.arplant.49.1.173. PMID 15012232. S2CID 6241264.
- ^ Madl P, Yip M (2004). "Literature Review of Caulerpa taxifolia". BUFUS-Info. 19 (31). Archived from the original on 8 October 2022. Retrieved 18 July 2017.
- ^ Orth RJ, Carruthers TJ, Dennison WC, Duarte CM, Fourqurean JW, Heck KL, et al. (2006). "A global crisis for seagrass ecosystems". BioScience. 56 (12): 987–996. doi:10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2. hdl:10261/88476. S2CID 4936412.
- ^ Froese, Rainer; Pauly, Daniel (eds.) (2009). "Phycodurus eques" in FishBase. July 2009 version.
- ^ Giri C, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, et al. (2011). "Status and distribution of mangrove forests of the world using earth observation satellite data". Global Ecology and Biogeography. 20 (1): 154–159. doi:10.1111/j.1466-8238.2010.00584.x.
- ^ Thomas N, Lucas R, Bunting P, Hardy A, Rosenqvist A, Simard M (2017). "Distribution and drivers of global mangrove forest change, 1996-2010". PLOS ONE. 12 (6): e0179302. Bibcode:2017PLoSO..1279302T. doi:10.1371/journal.pone.0179302. PMC 5464653. PMID 28594908.
- ^ Short FT, Frederick T (2003). World atlas of seagrasses. Berkeley, Calif.: University of California Press. p. 24. ISBN 978-0-520-24047-6. Archived from the original on 10 July 2019. Retrieved 10 July 2019.
- ^ Spalding M (2010). World atlas of mangroves. Routledge. doi:10.4324/9781849776608. ISBN 978-1-84977-660-8.
- ^ Lalli C, Parsons T (1993). Biological Oceanography: An Introduction. Butterworth-Heinemann. ISBN 0-7506-3384-0.
- ^ Lindsey R, Scott M, Simmon R (2010). "What are phytoplankton". NASA Earth Observatory.
- ^ Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (July 1998). "Primary production of the biosphere: integrating terrestrial and oceanic components". Science. 281 (5374): 237–40. Bibcode:1998Sci...281..237F. doi:10.1126/science.281.5374.237. PMID 9657713.
- ^ Rost B, Riebesell U (2004). "Coccolithophores and the biological pump: responses to environmental changes". Coccolithophores: From Molecular Processes to Global Impact. Springer. pp. 99–125. ISBN 978-3-662-06278-4.
- ^ Arsenieff L, Simon N, Rigaut-Jalabert F, Le Gall F, Chaffron S, Corre E, Com E, Bigeard E, Baudoux AC (2018). "First Viruses Infecting the Marine Diatom Guinardia delicatula". Frontiers in Microbiology. 9: 3235. doi:10.3389/fmicb.2018.03235. PMC 6334475. PMID 30687251.
- ^ Varea C, Aragón JL, Barrio RA (October 1999). "Turing patterns on a sphere". Physical Review E. 60 (4 Pt B): 4588–92. Bibcode:1999PhRvE..60.4588V. doi:10.1103/PhysRevE.60.4588. PMID 11970318.
- ^ Harvey EN (1952). Bioluminescence. Academic Press.
- ^ "Suggested Explanation for Glowing Seas--Including Currently Glowing California Seas". National Science Foundation. 18 October 2011.
- ^ Castro P, Huber ME (2010). Marine Biology (8th ed.). McGraw Hill. pp. 95. ISBN 978-0071113021.
- ^ Hastings JW (1996). "Chemistries and colors of bioluminescent reactions: a review". Gene. 173 (1 Spec No): 5–11. doi:10.1016/0378-1119(95)00676-1. PMID 8707056.
- ^ Haddock SH, Moline MA, Case JF (2009). "Bioluminescence in the sea". Annual Review of Marine Science. 2: 443–93. Bibcode:2010ARMS....2..443H. doi:10.1146/annurev-marine-120308-081028. PMID 21141672. S2CID 3872860.
- ^ "Carbon Cycling and Biosequestration". US Department of Energy Department of Energy Office of Science. 2008. p. 81.
Workshop report DOE/SC-108
- ^ Campbell M (22 June 2011). "The role of marine plankton in sequestration of carbon". EarthTimes. Retrieved 22 August 2014.
- ^ Roman J, McCarthy JJ (October 2010). "The whale pump: marine mammals enhance primary productivity in a coastal basin". PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007. e13255.
- ^ Brown JE (12 October 2010). "Whale poop pumps up ocean health". Science Daily. Retrieved 18 August 2014.
- ^ "Water, the Universal Solvent". USGS. Archived from the original on 9 July 2017. Retrieved 27 June 2017.
- ^ Brum JR, Morris JJ, Décima M, Stukel MR (2014). Chapter 2: Mortality in the oceans: Causes and consequences. Eco-DAS IX Symposium Proceedings. Association for the Sciences of Limnology and Oceanography. pp. 16–48. ISBN 978-0-9845591-3-8..
- ^ Reece JB (2013). Campbell Biology (10th ed.). Pearson. ISBN 978-0-321-77565-8.
- ^ Prentice IC (2001). "The carbon cycle and atmospheric carbon dioxide". Climate change 2001: the scientific basis: contribution of Working Group I to the Third Assessment Report of the Intergouvernmental Panel on Climate Change / Houghton, J.T. [edit.] Retrieved 31 May 2012.
- ^ Thomson CW (2014). Voyage of the Challenger: The Atlantic. Cambridge University Press. p. 235. ISBN 978-1-108-07475-9.
- ^ Hasle GR, Syvertsen EE, Steidinger, Tangen K (25 January 1996). "Marine Diatoms". In Tomas CR (ed.). Identifying Marine Diatoms and Dinoflagellates. Academic Press. pp. 5–385. ISBN 978-0-08-053441-1. Retrieved 13 November 2013.
- ^ Adl SM, Leander BS, Simpson AG, Archibald JM, Anderson OR, Bass D, et al. (August 2007). "Diversity, nomenclature, and taxonomy of protists". Systematic Biology. 56 (4): 684–9. doi:10.1080/10635150701494127. PMID 17661235.
- ^ Moheimani NR, Webb JP, Borowitzka MA (October 2012). "Bioremediation and other potential applications of coccolithophorid algae: a review". Algal Research. 1 (2): 120–33. doi:10.1016/j.algal.2012.06.002.
- ^ Taylor AR, Chrachri A, Wheeler G, Goddard H, Brownlee C (June 2011). "A voltage-gated H+ channel underlying pH homeostasis in calcifying coccolithophores". PLOS Biology. 9 (6): e1001085. doi:10.1371/journal.pbio.1001085. PMC 3119654. PMID 21713028.
- ^ Halpern BS, Frazier M, Afflerbach J, Lowndes JS, Micheli F, O'Hara C, et al. (August 2019). "Recent pace of change in human impact on the world's ocean". Scientific Reports. 9 (1): 11609. Bibcode:2019NatSR...911609H. doi:10.1038/s41598-019-47201-9. PMC 6691109. PMID 31406130.
- ^ "Human impacts on marine ecosystems". GEOMAR Helmholtz Centre for Ocean Research. Archived from the original on 31 October 2020. Retrieved 22 October 2019.
- ^ Rosing MT, Bird DK, Sleep NH, Bjerrum CJ (April 2010). "No climate paradox under the faint early Sun". Nature. 464 (7289): 744–7. Bibcode:2010Natur.464..744R. doi:10.1038/nature08955. PMID 20360739. S2CID 205220182.
- ^ a b Sahney S, Benton MJ, Ferry PA (August 2010). "Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land". Biology Letters. 6 (4): 544–7. doi:10.1098/rsbl.2009.1024. PMC 2936204. PMID 20106856.
- ^ McKinney ML (1997). "How do rare species avoid extinction? A paleontological view". The Biology of Rarity. pp. 110–29. doi:10.1007/978-94-011-5874-9_7. ISBN 978-94-010-6483-5.
- ^ Stearns BP, Stearns SC (1999). Watching, from the Edge of Extinction. New Haven, CT: Yale University Press. p. x. ISBN 978-0-300-08469-6.
- ^ Novacek MJ (8 November 2014). "Prehistory's Brilliant Future". The New York Times. New York. ISSN 0362-4331. Retrieved 25 December 2014.
- ^ Nee S (August 2004). "Extinction, slime, and bottoms". PLOS Biology. 2 (8): E272. doi:10.1371/journal.pbio.0020272. PMC 509315. PMID 15314670.
- ^ Ward PD (October 2006). "Impact from the deep". Scientific American. 295 (4): 64–71. Bibcode:2006SciAm.295d..64W. doi:10.1038/scientificamerican1006-64 (inactive 26 April 2024). PMID 16989482.
{{cite journal}}: CS1 maint: DOI inactive as of April 2024 (link) - ^ Marine Extinctions: Patterns and Processes - an overview. 2013. CIESM Monograph 45 [1]
- ^ a b c "Investigating Marine Life | Census of Marine Life". www.coml.org. Retrieved 27 December 2023.
- ^ "Exploration Tools: AUVs: NOAA Office of Ocean Exploration and Research". oceanexplorer.noaa.gov. Retrieved 27 December 2023.
- ^ "Deep-Towed Vehicles (DTVs) | Census of Marine Life". www.coml.org. Retrieved 27 December 2023.
- ^ "The Drone Revolution underwater". Advanced Navigation. Retrieved 27 December 2023.
Further reading
edit- Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D'Agrosa C, et al. (February 2008). "A global map of human impact on marine ecosystems". Science. 319 (5865): 948–52. Bibcode:2008Sci...319..948H. doi:10.1126/science.1149345. PMID 18276889. S2CID 26206024.
- Paleczny M, Hammill E, Karpouzi V, Pauly D (2015). "Population Trend of the World's Monitored Seabirds, 1950-2010". PLOS ONE. 10 (6): e0129342. Bibcode:2015PLoSO..1029342P. doi:10.1371/journal.pone.0129342. PMC 4461279. PMID 26058068.
- Ruppert EE, Fox RS, Barnes RD (2004). Invertebrate Zoology (7th ed.). Brooks / Cole. ISBN 978-0-03-025982-1.
- "After 60 million years of extreme living, seabirds are crashing". The Guardian. 22 September 2015.