The Stramenopiles, also called Heterokonts, are a clade of organisms distinguished by the presence of stiff tripartite external hairs. In most species, the hairs are attached to flagella, in some they are attached to other areas of the cellular surface, and in some they have been secondarily lost (in which case relatedness to stramenopile ancestors is evident from other shared cytological features or from genetic similarity). Stramenopiles represent one of the three major clades in the SAR supergroup, along with Alveolata and Rhizaria.
| Stramenopiles Temporal range: Late Mesoproterozoic-present,
| |
|---|---|
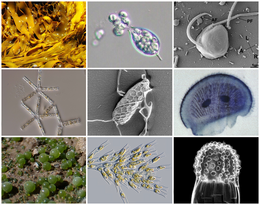 | |
| Diversity of stramenopiles | |
| Scientific classification | |
| Domain: | Eukaryota |
| Clade: | Diaphoretickes |
| Clade: | SAR |
| Clade: | Stramenopiles Patterson 1989[2] emend. Adl et al. 2005[3] |
| Phyla and subphyla[4] | |
| Diversity | |
| >100000 species[1] | |
| Synonyms | |
| |
Stramenopiles are eukaryotes; most are single-celled, but some are multicellular including some large seaweeds, the brown algae. The group includes a variety of algal protists, heterotrophic flagellates, opalines and closely related proteromonad flagellates (all endobionts in other organisms); the actinophryid heliozoa, and oomycetes. The tripartite hairs characteristic of the group have been lost in some of the included taxa – for example in most diatoms.
Many stramenopiles are unicellular flagellates, and most others produce flagellated cells at some point in their lifecycles, for instance as gametes or zoospores. Most flagellated heterokonts have two flagella; the anterior flagellum has one or two rows of stiff hairs or mastigonemes, and the posterior flagellum is without such embellishments, being smooth, usually shorter, or in a few cases not projecting from the cell.
The term 'heterokont' is used both as an adjective – indicating that a cell has two dissimilar flagella, and as the name of a taxon. The groups included in that taxon have however varied widely, creating the 'heterokont problem', now resolved by the definition of the stramenopiles.
History edit
The term 'stramenopile' was introduced by D. J. Patterson in 1989, defining a group that overlapped with the ambiguously defined heterokonts.[12][13] The name "stramenopile" has been discussed by J. C. David.[14]
The heterokont problem edit
The term 'heterokont' is used as both an adjective – indicating that a cell has two dissimilar flagella – and as the name of a taxon. The taxon 'Heterokontae' was introduced in 1899 by Alexander Luther for algae that are now considered the Xanthophyceae.[15] But the same term was used for other groupings of algae. For example, in 1956, Copeland[16] used it to include the xanthophytes (using the name Vaucheriacea), a group that included what became known as the chrysophytes, the silicoflagellates, and the hyphochytrids. Copeland also included the unrelated collar flagellates (as the choanoflagellates) in which he placed the bicosoecids. He also included the not-closely related haptophytes. The consequence of associating multiple concepts to the taxon 'heterokont' is that the meaning of 'heterokont' can only be made clear by making reference to its usage: Heterokontae sensu Luther 1899; Heterokontae sensu Copeland 1956, etc. This contextual clarification is rare, such that when the taxon name is used, it is unclear how it should be understood. The term 'Heterokont' has lost its usefulness in critical discussions about the identity, nature, character and relatedness of the group.[17] The term 'stramenopile' sought to identify a clade (monophyletic and holophyletic lineage) using the approach developed by transformed cladists of pointing to a defining innovative characteristic or apomorphy.[18]
Over time, the scope of application has changed, especially when in the 1970s ultrastructural studies revealed greater diversity among the algae with chromoplasts (chlorophylls a and c) than had previously been recognized. At the same time, a protistological perspective was replacing the 19th century one based on the division of unicellular eukaryotes into animals and plants. One consequence was that an array of heterotrophic organisms, many not previously considered as 'heterokonts', were seen as related to the 'core heterokonts' (those having anterior flagella with stiff hairs). Newly recognized relatives included the parasitic opalines, proteromonads, and actinophryid heliozoa. They joined other heterotrophic protists, such as bicosoecids, labyrinthulids, and oomycete fungi, that were included by some as heterokonts and excluded by others. Rather than continue to use a name whose meaning had changed over time and was hence ambiguous, the name 'stramenopile' was introduced to refer to the clade of protists that had tripartite stiff (usually flagellar) hairs and all their descendants. Molecular studies confirm that the genes that code for the proteins of these hairs are exclusive to stramenopiles.[19]
Characteristics edit


The presumed apomorphy of tripartite flagellar hairs in stramenopiles is well characterized. The basal part of the hair is flexible and inserts into the cell membrane; the second part is dominated by a long stiff tube (the 'straw' or 'stramen'); and finally the tube is tipped by many delicate hairs called mastigonemes.[20] The proteins that code for the mastigonemes appear to be exclusive to the stramenopile clade, and are present even in taxa (such as diatoms) that no longer have such hairs.[21]
Most stramenopiles have two flagella near the apex.[22] They are usually supported by four microtubule roots in a distinctive pattern. There is a transitional helix inside the flagellum where the beating axoneme with its distinctive geometric pattern of nine peripheral couplets around two central microtubules changes into the nine-triplet structure of the basal body.[23]
Plastids edit
Many stramenopiles have plastids which enable them to photosynthesise, using light to make their own food. Those plastids are coloured off-green, orange, golden or brown because of the presence of chlorophyll a, chlorophyll c, and fucoxanthin. This form of plastid is called a stramenochrome or chromoplast.[a] The most significant autotrophic stramenopiles are the brown algae (wracks and many other seaweeds), and the diatoms. The latter are among the most significant primary producers in marine and freshwater ecosystems.[24] Most molecular analyses suggest that the most basal stramenopiles lacked plastids and were accordingly colourless heterotrophs, feeding on other organisms. This implies that the stramenopiles arose as heterotrophs, diversified, and then some of them acquired chromoplasts. Some lineages (such as the axodine lineage that included the chromophytic pedinellids, colourless ciliophryids, and colourless actinophryid heliozoa) have secondarily reverted to heterotrophy.[25][26]
Ecology edit

Some stramenopiles are significant as autotrophs and as heterotrophs in natural ecosystems; others are parasitic. Blastocystis is a gastrointestinal parasite of humans;[27] opalines and proteromonads live in the intestines of cold-blooded vertebrates and have been described as parasitic;[28] oomycetes include some significant plant pathogens such as the cause of potato blight, Phytophthora infestans.[29] Diatoms are major contributors to global carbon cycles because they are the most important autotrophs in most marine habitats.[30] The brown algae, including familiar seaweeds like wrack and kelp, are major autotrophs of the intertidal and subtidal marine habitats.[31] Some of the bacterivorous stramenopiles, such as Cafeteria, are common and widespread consumers of bacteria, and thus play a major role in recycling carbon and nutrients within microbial food webs.[32][33]
Evolution edit
External edit
Stramenopiles are most closely related to Alveolates and Rhizaria, all of which have tubular mitochondrial cristae and collectively form the SAR supergroup, whose name is formed from their initials.[34][26][35] The ancestor of the SAR supergroup appears to have captured a unicellular photosynthetic red alga, and many Stramenopiles, as well as members of other SAR groups such as the Rhizaria, still have plastids which retain the double membrane of the red alga and a double membrane surrounding it, for a total of four membranes.[36] In addition, species of Telonemia, the sister group to SAR, exhibit heterokont flagella with tripartite mastigonemes, implying a more ancient origin of stramenopile characteristics.[37]
.jpg)
Internal edit
The following cladogram summarizes the evolutionary relationships between Stramenopiles. The phylogenetic relationships of Bigyra vary greatly from one analysis to the next: it has been recovered as either monophyletic[38][39] or paraphyletic. When paraphyletic, the branching order of the bigyran groups also varies: in some studies Sagenista is the most basal-branching clade,[38][40][41] while in others Opalozoa is the most basal.[42] Nonetheless, Platysulcea is consistently recovered as the sister clade to all other stramenopiles.[39][40] In addition, a flagellate species discovered in 2023, Kaonashia insperata, remains in an uncertain phylogenetic position, but more closely related to Gyrista than to other clades.[41]
| Stramenopiles | ||

Classification edit

The classification of the Stramenopiles according to Adl et al. (2019), with additions from newer research:[43] [4]
- Platysulcea Cavalier-Smith 2017[39]
- Bigyra Cavalier-Smith 1998, emend. 2006
- Opalozoa Cavalier Smith 1991, emend. 2006
- Nanomonadea Cavalier-Smith 2012
- Opalinata Wenyon 1926, emend. Cavalier-Smith 1997 [=Slopalinida Patterson 1985]
- Bicosoecida Grasse 1926, emend. Karpov 1998
- Sagenista Cavalier-Smith 1995
- Labyrinthulomycetes Dick 2001
- Pseudophyllomitidae Shiratori et al. 2016[40]
- Opalozoa Cavalier Smith 1991, emend. 2006
- Gyrista Cavalier-Smith 1998
- Bigyromonada Cavalier-Smith 1998
- Developea Karpov & Aleoshin 2016
- Pirsoniales Cavalier-Smith 1998, emend. 2006
- Pseudofungi Cavalier-Smith 1986
- Hyphochytriales Sparrow 1960
- Peronosporomycetes Dick 2001 [=Oomycetes Winter 1897, emend. Dick 1976]
- Actinophryidae Claus 1874, emend. Hartmann 1926
- Ochrophyta Cavalier-Smith 1986, emend. Cavalier-Smith & Chao 1996
- Chrysista Cavalier-Smith 1986
- Chrysoparadoxophyceae Wetherbee 2019[44]
- Chrysophyceae Pascher 1914
- Chloromorophyceae (nomen dubium)[45]
- Eustigmatophyceae Hibberd & Leedale 1971
- Olisthodiscophyceae Barcytė, Eikrem & M.Eliáš, 2021[46][47]
- Phaeophyceae Hansgirg 1886
- Phaeosacciophyceae R.A.Andersen, L.Graf & H.S.Yoon 2020[48]
- Phaeothamniophyceae Andersen & Bailey in Bailey et al. 1998
- Raphidophyceae Chadefaud 1950, emend. Silva 1980
- Schizocladiophyceae Henry, Okuda & Kawai, 2003
- Synchromophyceae S.Horn & C.Wilhelm 2007 [=Picophagea Cavalier-Smith 2006, emend. 2017]
- Xanthophyceae Allorge 1930, emend. Fritsch 1935 [Heterokontae Luther 1899; Heteromonadea Leedale 1983; Xanthophyta Hibberd 1990]
- Diatomista Derelle et al. 2016, emend. Cavalier-Smith 2017
- Bolidophyceae Guillou et al. 1999
- Diatomeae Dumortier 1821 [=Bacillariophyta Haeckel 1878]
- Dictyochophyceae Silva 1980
- Pelagophyceae Andersen & Saunders 1993
- Pinguiophyceae Kawachi et al. 2003
- Chrysista Cavalier-Smith 1986
- Bigyromonada Cavalier-Smith 1998
Notes edit
- ^ They are not called chloroplasts, the most common form of photosynthetic plastid. If used narrowly, a chloroplast is a plastid which contains chlorophyll B, as in green algae, some euglenids, and the land plants.
References edit
- ^ a b H.S. Yoon; R.A. Andersen; S.M. Boo; D. Bhattacharya (17 February 2009). "Stramnenopiles". Encyclopedia of Microbiology (Third Edition): 721–731. doi:10.1016/B978-012373944-5.00253-4. Retrieved 2 March 2024.
- ^ Patterson, D.J. (1989). "Stramenopiles: Chromophytes from a protistan perspective". In Green, J.C.; Leadbeater, B.S.C.; Diver, W.L. (eds.). The chromophyte algae: Problems and perspectives. Clarendon Press. ISBN 978-0198577133.
- ^ Sina M Adl; Alastair G B Simpson; Mark A Farmer; et al. (1 September 2005). "The new higher level classification of eukaryotes with emphasis on the taxonomy of protists". Journal of Eukaryotic Microbiology. 52 (5): 399–451. doi:10.1111/J.1550-7408.2005.00053.X. ISSN 1066-5234. PMID 16248873. Wikidata Q22065654.
- ^ a b Cavalier-Smith, Thomas (January 2018). "Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences". Protoplasma. 255 (1): 297–357. doi:10.1007/s00709-017-1147-3. PMC 5756292. PMID 28875267.
- ^ Vørs, N. (1993). "Marine heterotrophic amoebae, flagellates and heliozoa from Belize (Central America) and Tenerife". Journal of Eukaryotic Microbiology. 40 (3): 272–287. doi:10.1111/j.1550-7408.1993.tb04917.x. S2CID 221852241.
- ^ David, J.C. (2002). "A preliminary catalogue of the names of fungi above the rank of order". Constancea. 83: 1–30.
- ^ Cavalier-Smith, T. (1999). "The kingdom Chromista, origin and systematics". In Round, F.E.; Chapman, D.J. (eds.). Progress in Phycological Research. Vol. 4. Elsevier. pp. 309–347. ISBN 978-0-948737-00-8.
- ^ van den Hoek, C.; Mann, D.G.; Jahns, H.M. (1995). Algae An Introduction to Phycology. Cambridge University Press. ISBN 978-0-521-30419-1.
- ^ Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. (1996). Introductory Mycology (4th ed.). Wiley. ISBN 978-0471522294.
- ^ Dick, M.W. (2013). Straminipilous Fungi: Systematics of the Peronosporomycetes Including Accounts of the Marine Straminipilous Protists, the Plasmodiophorids and Similar Organisms. Springer. ISBN 978-94-015-9733-3.
- ^ "Stramenipila M.W. Dick (2001)". MycoBank. International Mycological Association.
- ^ Patterson, D. J. (1989). "Stramenopiles: chromophytes from a protistological perspective". In Green, J. C.; Leadbeater, B. S. C.; Diver, W. L. (eds.). The chromophyte algae: problems and perspectives. Oxford: Clarendon Press. pp. 357–379.
- ^ Patterson, David J. (1999). "The Diversity of Eukaryotes". The American Naturalist. 154 (S4): S96–S124. doi:10.1086/303287. PMID 10527921. S2CID 4367158.
- ^ David, J. C. (2002). "A preliminary catalogue of the names of fungi above the rank of order". Constancea (83): 1–30.
- ^ Luther, Alexander F. (1899). Über Chlorosaccus eine neue Gattung der Süsswasseralgen nebst einiger Bemerkungen zur Systematik verwandter Algen [About Chlorosaccus a new genus of freshwater algae together with some comments on the systematics of related algae] (in German). Stockholm: Norstedt. pp. 1–22.
- ^ Copeland, H. F. (1956). The Classification of Lower Organisms. Palo Alto, California: Pacific Books.
- ^ Blackwell, W. H. (2009). "Chromista revisited: A dilemma of overlapping putative kingdoms, and the attempted application of the botanical code of nomenclature" (PDF). Phytologia. 91 (2).
- ^ Patterson, Colin (1982). "Morphological characters and homology". In Joysey, Kenneth A.; Friday, A. E. (eds.). Problems in Phylogenetic Reconstruction. Systematics Association Special Volume 21. London: Academic Press. ISBN 978-0-1239-1250-3.
- ^ Hee, Wei Yih; Blackman, Leila M.; Hardham, Adrienne R. (2019). "Characterisation of Stramenopile-specific mastigoneme proteins in Phytophthora parasitica". Protoplasma. 256 (2): 521–535. doi:10.1007/s00709-018-1314-1. PMID 30302550. S2CID 52947780.
- ^ Bouck, G. Benjamin (1 August 1971). "The structure, origin, and composition of the tubular mastigonemes of the Ochromonas flagellum". Journal of Cell Biology. 50 (2): 362–384. doi:10.1083/jcb.50.2.362. PMC 2108286. PMID 5123323.
- ^ Blackman, Leila M.; Arikawa, Mikihiko; Yamada, Shuhei; Suzaki, Toshinobu; Hardham, Adrienne R. (2011). "Identification of a Mastigoneme Protein from Phytophthora nicotianae". Protist. 162 (1): 100–114. doi:10.1016/j.protis.2010.01.005. PMID 20663714.
- ^ Yoon, H.S.; Andersen, R.A.; Boo, S.M.; Bhattacharya, D. (2009). "Stramenopiles". Encyclopedia of Microbiology. Elsevier. pp. 721–731. doi:10.1016/b978-012373944-5.00253-4. ISBN 9780123739445.
- ^ Fu, Gang; Nagasato, Chikako; Oka, Seiko; Cock, J. Mark; Motomura, Taizo (2014). "Proteomics Analysis of Heterogeneous Flagella in Brown Algae (Stramenopiles)" (PDF). Protist. 165 (5): 662–675. doi:10.1016/j.protis.2014.07.007. PMID 25150613. S2CID 7936118.
- ^ Leipe, D. D.; Wainright, P. O.; Gunderson, J. H.; et al. (1994). "The stramenopiles from a molecular perspective: 16S-like rRNA sequences from Labyrinthuloides minuta and Cafeteria roenbergensis". Phycologia. 33 (5): 369–377. doi:10.2216/i0031-8884-33-5-369.1.
- ^ Leyland, Ben; Leu, Stefan; Boussiba, Sammy (2017). "Are Thraustochytrids algae?". Fungal Biology. 121 (10): 835–840. doi:10.1016/j.funbio.2017.07.006. PMID 28889907.
- ^ a b Derelle, Romain; López-García, Purificación; Timpano, Hélène; Moreira, David (10 August 2016). "A Phylogenomic Framework to Study the Diversity and Evolution of Stramenopiles (=Heterokonts)". Molecular Biology and Evolution. 33 (11): 2890–2898. doi:10.1093/molbev/msw168. PMC 5482393. PMID 27512113.
- ^ Roberts, Tamalee; Stark, Damien; Harkness, John; Ellis, John (2014). "Update on the pathogenic potential and treatment options for Blastocystis sp". Gut Pathogens. 6 (1): 17. doi:10.1186/1757-4749-6-17. PMC 4039988. PMID 24883113.
- ^ Olsen, O. Wilford (1986). Animal Parasites : their life cycles and ecology. New York: Dover. pp. 56, 74–75. ISBN 0-486-65126-6. OCLC 13123309.
- ^ Nowicki, Marcin; et al. (17 August 2011). "Potato and tomato late blight caused by Phytophthora infestans: An overview of pathology and resistance breeding". Plant Disease. 96 (1). American Phytopathological Society: 4–17. doi:10.1094/PDIS-05-11-0458. PMID 30731850.
- ^ Yool, Andrew; Tyrrell, Toby (2003). "Role of diatoms in regulating the ocean's silicon cycle". Global Biogeochemical Cycles. 17 (4): n/a. Bibcode:2003GBioC..17.1103Y. CiteSeerX 10.1.1.394.3912. doi:10.1029/2002GB002018. S2CID 16849373.
- ^ Cock, J. Mark; Peters, Akira F.; Coelho, Susana M. (2011-08-09). "Brown algae". Current Biology. 21 (15): R573–R575. doi:10.1016/j.cub.2011.05.006. PMID 21820616.
- ^ Guiry, Wendy (7 April 2011). "Cafeteria T.Fenchel & D.J.Patterson 1988". AlgaeBase. Retrieved 17 March 2023.
- ^ Fenchel, T.; Patterson, D. J. (1988). "Cafeteria roenbergensis nov. gen., nov. sp., a heterotrophic microflagellate from marine plankton". Marine Microbial Food Webs. 3: 9–19.
- ^ Krylov, M. V.; Dobrovolskii, A. A.; Issi, I. V.; Michaelevich, B. I.; Podlipaev, S. A.; Reshetnyak, V. V.; Seravin, L. N.; et al. 1980. New concepts for the system of unicellular organisms. Trudy Zoologischkei Institut Akademiya Nayuk, SSSR 94:122–132.
- ^ Burki, F.; Shalchian-Tabrizi; Pawlowski, J. (August 2008). "Phylogenomics reveals a new 'megagroup' including most photosynthetic eukaryotes". Biology Letters. 4 (4): 366–369. doi:10.1098/rsbl.2008.0224. PMC 2610160. PMID 18522922.
- ^ Oborník, Miroslav; Lukeš, Julius (2013). "Cell Biology of Chromerids". International Review of Cell and Molecular Biology. Vol. 306. Elsevier. pp. 333–369. doi:10.1016/b978-0-12-407694-5.00008-0. ISBN 9780124076945. PMID 24016529.
- ^ Tikhonenkov, Denis V.; Jamy, Mahwash; Borodina, Anastasia S.; Belyaev, Artem O.; Zagumyonnyi, Dmitry G.; Prokina, Kristina I.; Mylnikov, Alexander P.; Burki, Fabien; Karpov, Sergey A. (2022). "On the origin of TSAR: morphology, diversity and phylogeny of Telonemia". Open Biology. 12 (3). The Royal Society. doi:10.1098/rsob.210325. ISSN 2046-2441. PMC 8924772. PMID 35291881.
- ^ a b Cavalier-Smith, Thomas; Scoble, Josephine Margaret (2013). "Phylogeny of Heterokonta: Incisomonas marina, a uniciliate gliding opalozoan related to Solenicola (Nanomonadea), and evidence that Actinophryida evolved from raphidophytes". European Journal of Protistology. 49 (3): 328–353. doi:10.1016/j.ejop.2012.09.002. PMID 23219323.
- ^ a b c Thakur, Rabindra; Shiratori, Takashi; Ishida, Ken-ichiro (2019). "Taxon-rich Multigene Phylogenetic Analyses Resolve the Phylogenetic Relationship Among Deep-branching Stramenopiles". Protist. 170 (5): 125682. doi:10.1016/j.protis.2019.125682. ISSN 1434-4610. PMID 31568885. S2CID 202865459.
- ^ a b c Shiratori, Takashi; Thakur, Rabindra; Ishida, Ken-ichiro (2017). "Pseudophyllomitus vesiculosus (Larsen and Patterson 1990) Lee, 2002, a Poorly Studied Phagotrophic Biflagellate is the First Characterized Member of Stramenopile Environmental Clade MAST-6". Protist. 168 (4): 439–451. doi:10.1016/j.protis.2017.06.004. ISSN 1434-4610. PMID 28822908.
- ^ a b Elizabeth J. Weston; Yana Eglit; Alastair G. B. Simpson (6 October 2023). "Kaonashia insperata gen. et sp. nov., a eukaryotrophic flagellate, represents a novel major lineage of heterotrophic stramenopiles". Journal of Eukaryotic Microbiology. doi:10.1111/JEU.13003. ISSN 1066-5234. Wikidata Q123562228.
- ^ Anna Cho; Denis V. Tikhonenkov; Gordon Lax; Kristina I. Prokina; Patrick J. Keeling (10 November 2023). "Phylogenomic position of genetically diverse phagotrophic stramenopile flagellates in the sediment-associated MAST-6 lineage and a potentially halotolerant placididean". Molecular Phylogenetics and Evolution. 190: 107964. doi:10.1016/J.YMPEV.2023.107964. ISSN 1055-7903. PMID 37951557. Wikidata Q124719861.
- ^ Adl SM, Bass D, Lane CE, Lukeš J, Schoch CL, Smirnov A, Agatha S, Berney C, Brown MW, Burki F, Cárdenas P, Čepička I, Chistyakova L, del Campo J, Dunthorn M, Edvardsen B, Eglit Y, Guillou L, Hampl V, Heiss AA, Hoppenrath M, James TY, Karnkowska A, Karpov S, Kim E, Kolisko M, Kudryavtsev A, Lahr DJG, Lara E, Le Gall L, Lynn DH, Mann DG, Massana R, Mitchell EAD, Morrow C, Park JS, Pawlowski JW, Powell MJ, Richter DJ, Rueckert S, Shadwick L, Shimano S, Spiegel FW, Torruella G, Youssef N, Zlatogursky V, Zhang Q (2019). "Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes". Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi:10.1111/jeu.12691. PMC 6492006. PMID 30257078.
- ^ Wetherbee R, Jackson CJ, Repetti SI, Clementson LA, Costa JF, van de Meene A, Crawford S, Verbruggen H (April 2019). "The golden paradox - a new heterokont lineage with chloroplasts surrounded by two membranes". J Phycol. 55 (2): 257–278. doi:10.1111/jpy.12822. hdl:11343/233613. PMID 30536815. S2CID 54477112.
- ^ Medlin LK, Desdevises Y (2018). "Sequence analysis confirms a new algal class". Vie et Milieu/Life & Environment.
- ^ Graf, Louis; Yoon, Hwan Su (21 July 2021). "Olisthodiscophyceae, the 17th heterokont algal class". Journal of Phycology. 57 (4): 1091–1093. doi:10.1111/jpy.13184. PMID 34289104. S2CID 236175098.
- ^ Barcytė, Dovilė; Eikrem, Wenche; Engesmo, Anette; Seoane, Sergio; Wohlmann, Jens; Horák, Aleš; Yurchenko, Tatiana; Eliáš, Marek (2 March 2021). "Olisthodiscus represents a new class of Ochrophyta". Journal of Phycology. 57 (4): 1094–1118. doi:10.1111/jpy.13155. hdl:10852/86515. PMID 33655496. S2CID 232101186.
- ^ Graf L, Yang EC, Han KY, Küpper FC, Benes KM, Oyadomari JK, Herbert RJH, Verbruggen H, Wetherbee R, Andersen RA, Yoon HS (December 2020). "Multigene Phylogeny, Morphological Observation and Re-examination of the Literature Lead to the Description of the Phaeosacciophyceae Classis Nova and Four New Species of the Heterokontophyta SI Clade". Protist. 171 (6): 125781. doi:10.1016/j.protis.2020.125781. PMID 33278705. S2CID 227315556.