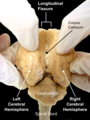
The vertebrate cerebrum (brain) is formed by two cerebral hemispheres that are separated by a groove, the longitudinal fissure. The brain can thus be described as being divided into left and right cerebral hemispheres. Each of these hemispheres has an outer layer of grey matter, the cerebral cortex, that is supported by an inner layer of white matter. In eutherian (placental) mammals, the hemispheres are linked by the corpus callosum, a very large bundle of nerve fibers. Smaller commissures, including the anterior commissure, the posterior commissure and the fornix, also join the hemispheres and these are also present in other vertebrates. These commissures transfer information between the two hemispheres to coordinate localized functions.
| Cerebral hemisphere | |
|---|---|
 Human brain seen from front. | |
 Right cerebral hemisphere Left cerebral hemisphere | |
| Details | |
| Identifiers | |
| Latin | hemisphaerium cerebri |
| NeuroNames | 241 |
| NeuroLex ID | birnlex_1796 |
| TA98 | A14.1.09.002 |
| TA2 | 5418 |
| FMA | 61817 |
| Anatomical terms of neuroanatomy | |
There are three known poles of the cerebral hemispheres: the occipital pole, the frontal pole, and the temporal pole.
The central sulcus is a prominent fissure which separates the parietal lobe from the frontal lobe and the primary motor cortex from the primary somatosensory cortex.
Macroscopically the hemispheres are roughly mirror images of each other, with only subtle differences, such as the Yakovlevian torque seen in the human brain, which is a slight warping of the right side, bringing it just forward of the left side. On a microscopic level, the cytoarchitecture of the cerebral cortex, shows the functions of cells, quantities of neurotransmitter levels and receptor subtypes to be markedly asymmetrical between the hemispheres.[1][2] However, while some of these hemispheric distribution differences are consistent across human beings, or even across some species, many observable distribution differences vary from individual to individual within a given species.
Structure edit
Each cerebral hemisphere has an outer layer of cerebral cortex which is of grey matter and in the interior of the cerebral hemispheres is an inner layer or core of white matter known as the centrum semiovale.[3] The interior portion of the hemispheres of the cerebrum includes the lateral ventricles, the basal ganglia, and the white matter.[4]
Poles edit
_-_inferiror_view.png)
There are three poles of the cerebrum: the occipital pole, the frontal pole, and the temporal pole. The occipital pole is the posterior end of each occipital lobe in each hemisphere. It is more pointed than the rounder frontal pole. The frontal pole is at the frontmost part of the frontal lobe in each hemisphere, and is more rounded than the occipital pole. The temporal pole is located between the frontal and occipital poles, and sits in the anterior part of middle cranial fossa in each temporal lobe.[5]
Composition edit
If the upper part of either hemisphere is removed, at a level about 1.25 cm above the corpus callosum, the central white matter will be exposed as an oval-shaped area, the centrum semiovale, surrounded by a narrow convoluted margin of gray substance, and studded with numerous minute red dots (puncta vasculosa), produced by the escape of blood from divided blood vessels.[citation needed]
If the remaining portions of the hemispheres be slightly drawn apart a broad band of white substance, the corpus callosum, will be observed, connecting them at the bottom of the longitudinal fissure; the margins of the hemispheres which overlap the corpus callosum are called the labia cerebri.[6]
Each labium is part of the cingulate gyrus already described; and the groove between it and the upper surface of the corpus callosum is termed the callosal sulcus.
If the hemispheres are sliced off to a level with the upper surface of the corpus callosum, the white substance of that structure will be seen connecting the two hemispheres.
The large expanse of medullary matter now exposed, surrounded by the convoluted margin of gray substance, is called the centrum semiovale. The blood supply to the centrum semiovale is from the superficial middle cerebral artery.[3] The cortical branches of this artery descend to provide blood to the centrum semiovale.[7]
Development edit
The cerebral hemispheres are derived from the telencephalon. They arise five weeks after conception as bilateral invaginations of the walls. The hemispheres grow round in a C-shape and then back again, pulling all structures internal to the hemispheres (such as the ventricles) with them. The intraventricular foramina (also called the foramina of Monro) allows communication with the lateral ventricles. The choroid plexus is formed from ependymal cells and vascular mesenchyme.[citation needed]
Function edit
Hemisphere lateralization edit
Broad generalizations are often made in popular psychology about certain functions (e.g. logic, creativity) being lateralized, that is, located in the right or left side of the brain. These claims are often inaccurate, as most brain functions are actually distributed across both hemispheres. Most scientific evidence for asymmetry relates to low-level perceptual functions rather than the higher-level functions popularly discussed (e.g. subconscious processing of grammar, not "logical thinking" in general).[8][9] In addition to this lateralization of some functions, the low-level representations also tend to represent the contralateral side of the body.
The best example of an established lateralization is that of Broca's and Wernicke's Areas (language) where both are often found exclusively on the left hemisphere. These areas frequently correspond to handedness however, meaning the localization of these areas is regularly found on the hemisphere opposite to the dominant hand. Function lateralization, such as semantics, intonation, accentuation, and prosody, has since been called into question and largely been found to have a neuronal basis in both hemispheres.[10][11]

Perceptual information is processed in both hemispheres, but is laterally partitioned: information from each side of the body is sent to the opposite hemisphere (visual information is partitioned somewhat differently, but still lateralized). Similarly, motor control signals sent out to the body also come from the hemisphere on the opposite side. Thus, hand preference (which hand someone prefers to use) is also related to hemisphere lateralization.[citation needed]
In some aspects, the hemispheres are asymmetrical; the right side is slightly bigger. There are higher levels of the neurotransmitter norepinephrine on the right and higher levels of dopamine on the left. The right hemisphere is more sensitive to testosterone. There is more white matter (longer axons) on the right and more grey matter (cell bodies) on the left.[12]
Linear reasoning functions of language such as grammar and word production are often lateralized to the left hemisphere of the brain. In contrast, holistic reasoning functions of language such as intonation and emphasis are often lateralized to the right hemisphere of the brain. Other integrative functions such as intuitive or heuristic arithmetic, binaural sound localization, etc. seem to be more bilaterally controlled.[13]
Clinical significance edit
Infarcts of the centrum ovale can occur.[3]
As a treatment for epilepsy the corpus callosum may be severed to cut the major connection between the hemispheres in a procedure known as a corpus callosotomy.
A hemispherectomy is the removal or disabling of one of the hemispheres of the brain. This is a rare procedure used in some extreme cases of seizures which are unresponsive to other treatments.
Additional images edit
-
 The sheep brain seen from the back. Opening longitudinal fissure which separates left and right cerebral hemispheres.
The sheep brain seen from the back. Opening longitudinal fissure which separates left and right cerebral hemispheres. -
 Lateral surface. (The frontal pole is approximately at 10, the occipital pole is approximately at 17, and the temporal pole is approximately at 38.)
Lateral surface. (The frontal pole is approximately at 10, the occipital pole is approximately at 17, and the temporal pole is approximately at 38.)


References edit
- ^ Anderson B, Rutledge V (December 1996). "Age and hemisphere effects on dendritic structure". Brain. 119 (6): 1983–1990. doi:10.1093/brain/119.6.1983. PMID 9010002.
- ^ Hutsler J, Galuske RA (August 2003). "Hemispheric asymmetries in cerebral cortical networks". Trends in Neurosciences. 26 (8): 429–435. CiteSeerX 10.1.1.133.2360. doi:10.1016/S0166-2236(03)00198-X. PMID 12900174. S2CID 15968665.
- ^ a b c Bogousslavsky J, Regli F (October 1992). "Centrum ovale infarcts: subcortical infarction in the superficial territory of the middle cerebral artery". Neurology. 42 (10): 1992–1998. doi:10.1212/wnl.42.10.1992. PMID 1340771. S2CID 219195107.
- ^ Snell RS (2009). Clinical Neuroanatomy for Medical Students. Hagerstwon, MD: Lippincott Williams & Wilkins. p. 262. ISBN 978-0-7817-9427-5.
- ^ Singh V (2014). "Cerebrum". Textbook of Anatomy Head, Neck, and Brain. Vol. III. India: Elsevier. p. 389. ISBN 978-81-312-3727-4.
- ^ Corsini R (5 December 2016). Labia cerebri. ISBN 9781317705703. Retrieved 13 August 2019.
- ^ Lee PH, Oh SH, Bang OY, Joo IS, Huh K (December 2005). "Pathogenesis of deep white matter medullary infarcts: a diffusion weighted magnetic resonance imaging study". Journal of Neurology, Neurosurgery, and Psychiatry. 76 (12): 1659–1663. doi:10.1136/jnnp.2005.066860. PMC 1739473. PMID 16291890.
- ^ Westen D, Burton LJ, Kowalski R (2006). Psychology (Australian and New Zealand ed.). John Wiley & Sons Australia, Ltd. p. 107. ISBN 978-0-470-80552-7.
- ^ "Neuromyth 6: The left brain/ right brain myth". Organisation for Economic Co-operation and Development (OECD). Retrieved 15 October 2011.
- ^ Weiss PH, Ubben SD, Kaesberg S, Kalbe E, Kessler J, Liebig T, Fink GR (January 2016). "Where language meets meaningful action: a combined behavior and lesion analysis of aphasia and apraxia". Brain Structure & Function. 221 (1): 563–576. doi:10.1007/s00429-014-0925-3. PMID 25352157. S2CID 16060074.
- ^ Riès SK, Dronkers NF, Knight RT (April 2016). "Choosing words: left hemisphere, right hemisphere, or both? Perspective on the lateralization of word retrieval". Annals of the New York Academy of Sciences. 1369 (1): 111–31. Bibcode:2016NYASA1369..111R. doi:10.1111/nyas.12993. PMC 4874870. PMID 26766393.
- ^ Carter R (1999). Mapping the mind. Berkeley, CA.: University of California Press. ISBN 978-0-520-22461-2.
- ^ Dehaene S, Spelke E, Pinel P, Stanescu R, Tsivkin S (May 1999). "Sources of mathematical thinking: behavioral and brain-imaging evidence". Science. 284 (5416): 970–974. Bibcode:1999Sci...284..970D. doi:10.1126/science.284.5416.970. PMID 10320379.